Катоды медные
Назначение и описание
Катоды медные выпускаются по ГОСТ 546, химический состав соответствует ГОСТ 859.
Предназначены для изготовления проволоки, шин, катанки, медных электролитических порошков.
Химический состав
| Элемент | Массовая доля, % | |||
| М1к | М0к | М00к | ||
| Cu | Медь, не менее | 99,95 | 99,97 | –* |
| Примеси, не более: | ||||
| Bi | Висмут | 0,0005 | 0,0003** | |
| Sn | Олово | 0,002 | 0,001 | – |
| Ni | Никель | 0,002*** | ||
| Fe | Железо | 0,003 | 0,001 | |
| Zn | Цинк | – | ||
| As | Мышьяк | 0,002 | 0,0005 | |
| P | Фосфор | 0,0015**** | ||
| Sb | Сурьма | 0,0004 | ||
| Pb | Свинец | 0,003 | 0,0005 | |
| Ag | Серебро | 0,002 | 0,002 | |
| S | Cера | 0,004 | 0,0015 | |
| O2 | Кислород | 0,02 | 0,015 | – |
* Знак «–» означает, что данный элемент определяется, но не нормируется.

** Сумма висмута, селена и теллура. Максимальное содержание каждого элемента не должно превышать 0,0002 %.
*** Сумма никеля, цинка, кремния, олова, железа, кобальта.
**** Сумма фосфора, хрома, марганца, мышьяка, кадмия, сурьмы Упаковка и транспортировка
Катоды формируют в пакеты массой не более 1500 кг и высотой не более 500 мм в соответствии с требованиями ГОСТ 21399. Транспортируют всеми видами крытых транспортных средств.
Гарантийный срок хранения
Срок хранения не ограничен.
Качество
Система менеджмента качества АО «Уралэлектромедь» сертифицирована компанией LRQA на соответствие требованиям ISO 9001:2008.
Системы экологического менеджмента, управления охраной труда и промышленной безопасностью, энергетического менеджмента сертифицированы Ассоциацией по сертификации Русский Регистр на соответствие требованиям международных стандартов ISO 14001:2004, OHSAS 18001:2007, ISO 50001:2011.

Катоды медные АО «Уралэлектромедь» зарегистриров
аны на Лондонской бирже металлов: в 2010 г. под брендом UMMC — отделения электролиза ЦЭМ, в 2013 г. под брендом UMMC II — отделения безосновной технологии нового ЦЭМ.
Продукция зарегистрирована в соответствии с европейским регламентом о безопасности химической продукции REACH.
По вопросам приобретения продукции:
- Начальник управления сбыта ОАО «УГМК» Храмов Олег Валерьевич +7(34368)9-68-13
- Заместитель начальника отдела сбыта ОАО «УГМК» Плотникова Яна Евгеньевна +7(34368)9-69-13
Медь — Сталь — Медь « Калужский завод по обработке цветных металлов
ТУ 1881-003-75479902-2005 Ленты многослойные марки МСтМ. Технические условияОписаниеМногослойная биметаллическая лента марки МСтМ, изготавливается из низкоуглеродистой стали, плакированной с двух сторон медью.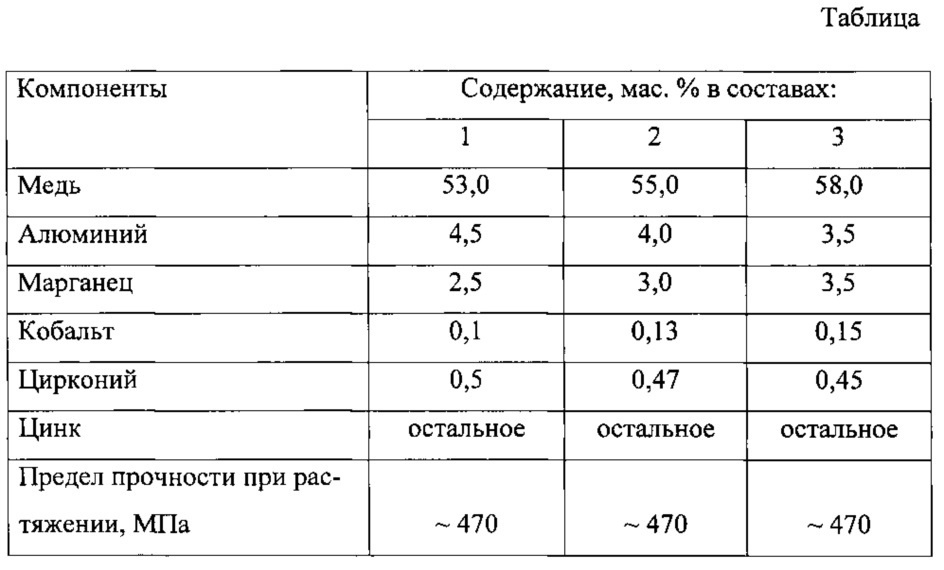
- Основа – низкоуглеродистая качественная сталь марки 08Ю с химическим составом по ГОСТ 9045.
- Покрытие – медь марки М1 с химическим составом по ГОСТ 859.
Толщина плакирующего слоя h2 и h3 является одинаковой для обеих сторон и составляет от 4 до 6% от толщины готовой ленты. По требованию Потребителя толщина плакирующего слоя может быть до 10% на сторону.
Классификация и сортаментПо состоянию материала:
- мягкое – М
- дрессированное — Д
- полутвёрдое – П
- твёрдое – Т
По виду кромок:
- с обрезной кромкой – О
- без обрезной кромки – Н
| Сорт | Толщина H, мм | Ширина B, мм | Допуск по ширине, мм | Длина, мм | Внутренний диаметр рулона Øвнутр.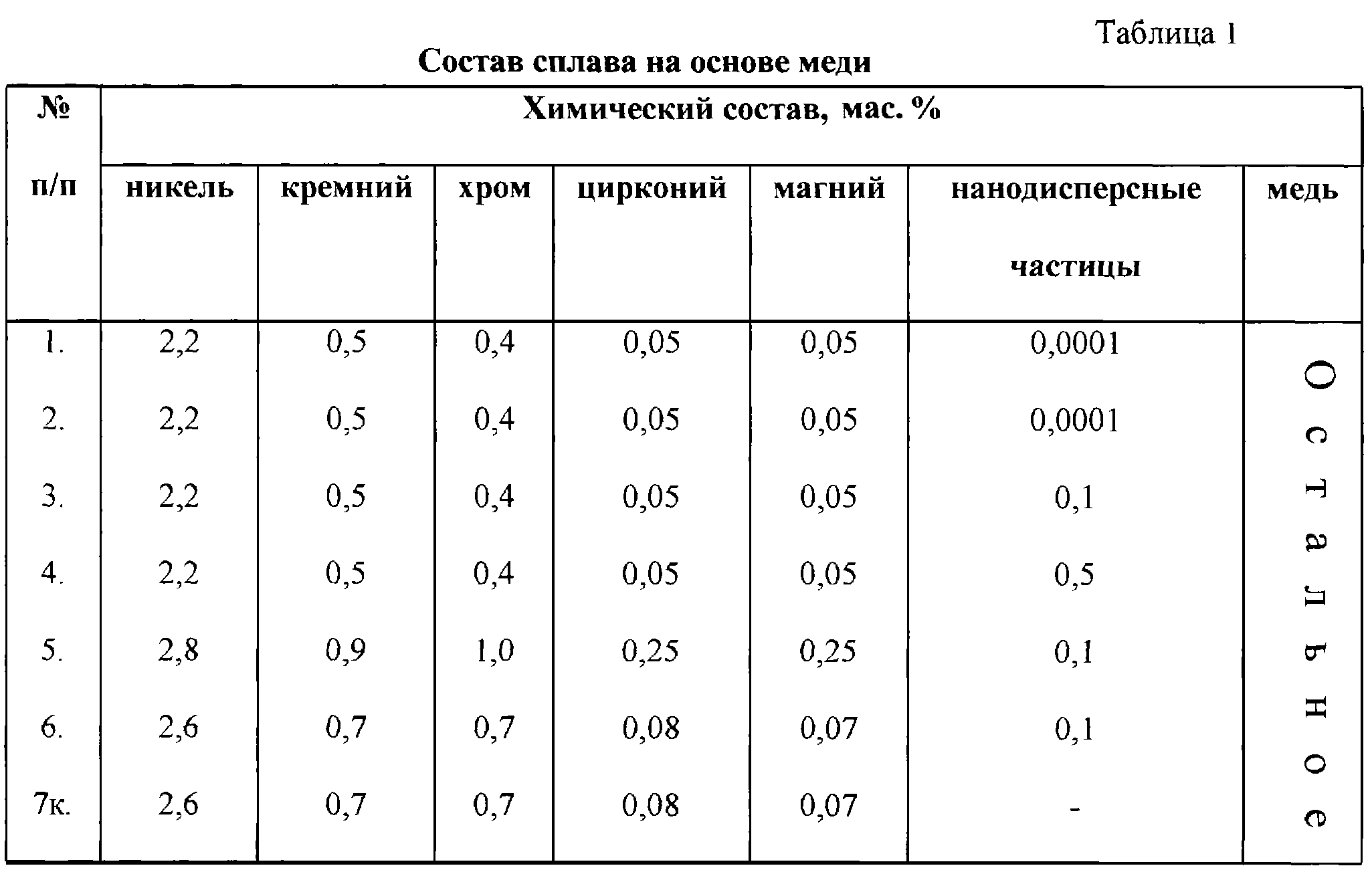 , мм , мм | Масса рулона, кг | |
Лента | 0,10 – 0,80 | от (±0,005)до (±0,03) | 6 – 60 | (±0,2) | немерная | 80 / 100 / 200 /300 / 400 | 10 – 100 |
60 – 220 | от (±0,2)до (±0,3) | 50 – 500 | |||||
0,81 – 1,50 | от (±0,03)до (±0,035) | 20 – 60 | (±0,2) | немерная | 300 / 400 / 500 | 10 – 100 | |
60 – 220 | от (±0,2)до (±0,3) | 50 – 500 | |||||
Полоса | 0,50 – 1,50 | от (±0,02)до (±0,035) | 50 – 220 | от (±0,2)до (±0,3) | 1000 – 2000 | — | — |
Для лент с необрезной кромкой допуск по ширине составляет (±1,0) мм.
Возможно изготовление других типоразмеров по согласованному допуску.
| Состояние ленты | Временное сопротивление разрыву σB, МПа | Относительное удлинениеδ, % |
Мягкое (М) | 250 ÷ 380 | ≥32 |
Дрессированное (Д) | 280 ÷ 400 | ≥26 |
Полутвёрдое (П) | 290 ÷ 430 | ≥10 |
Твёрдое (Т) | 370 ÷ 510 | ≥4 |
Для производства изделий электротехнической и автомобильной промышленности.
Русская Медная Компания — официальный сайт
#ПОМОЩЬУЖЕЗДЕСЬ
РМК помогает бороться с вирусом и преодолевать последствия пандемии
Системообразующее
предприятие
РМК сохраняет стабильность производства в условиях пандемии COVID-19
Металл будущего
Наша специализация — добыча и производство меди, ключевого ресурса для развития «умных» высокотехнологичных отраслей.
Устойчивое развитие
Решая бизнес-задачи, мы стараемся учитывать долгосрочные интересы общества: экономические, экологические и социальные.
Вертикальная интеграция
Предприятия РМК осуществляют полный цикл производства — от добычи и обогащения руды до выпуска и реализации готовой продукции.
Экологическая безопасность и охрана труда
Мы гарантируем максимальную безопасность и экологичность своего производства.
Инновационный подход
Мы изменяем традиционные представления о горной добыче и металлургическом производстве, внедряя инновации и совершенствуя подходы.
Люди — главный актив
РМК уделяет огромное внимание обеспечению благополучия, профессиональному развитию, социальной поддержке своих сотрудников и их семей.
Почему электрические кабеля делают из меди: особенности производства медного кабеля
Метод получения: | Массовая доля, % | |||||||
Медь Cu | Медь+Серебро Cu+Ag | примеси, не более | ||||||
не менее | Висмут Bi | Железо Fe | Никель Ni | Цинк Zn | Олово Sn | |||
М 00б | Переплавка катодов в восстановительной инертной атмосфере или вакууме. | 99,99 | – | 0,0005 | 0,001 | 0,001 | 0,001 | 0,001 |
М 0б | – | 99,97 | 0,001 | 0,004 | 0,002 | 0,003 | 0,002 | |
М 1б | – | 99,95 | 0,001 | 0,004 | 0,002 | 0,003 | 0,002 | |
М 00 | Переплавка катодов, полученных гальваническим способом. | 99,96 | – | 0,0005 | 0,001 | 0,001 | 0,001 | 0,001 |
М 0 | – | 99,93 | 0,0005 | 0,004 | 0,002 | 0,003 | 0,001 | |
М 1 | – | 99,90 | 0,001 | 0,005 | 0,002 | 0,004 | 0,002 | |
М 1р | Переплавка катодов и лома меди с раскислением фосфором. | – | 99,90 | 0,001 | 0,005 | 0,002 | 0,005 | 0,002 |
М 1ф | – | 99,90 | 0,001 | 0,005 | 0,002 | 0,005 | 0,002 | |
М 2р | – | 99,70 | 0,002 | 0,05 | 0,2 | – | 0,05 | |
М 3р | – | 99,50 | 0,003 | 0,05 | 0,2 | – | 0,05 | |
М 2 | Огневое рафинирование и переплавка отходов и лома меди. | – | 99,70 | 0,002 | 0,05 | 0,2 | – | 0,05 |
М 3 | – | 99,50 | 0,003 | 0,05 | 0,2 | – | 0,05 | |
М00к | Переплавка катодов, полученных гальваническим способом. | 99,98 | — | 0,0002 | 0,001 | — | — | — |


Марки и химический состав технической меди:
Химический состав марок меди стандартизируется в ГОСТ 859-2001
Разработан в Минске, внесен правительством РФ, принят всеми странами СНГ, в том числе и Украиной.
ГОСТ 859-2001 | EN, DIN |
М00 | Cu — OFF |
М0 | Cu-PHC, OF-Cu |
М1 | Cu-OF, Cu-OF1 |
M1 | SE-Cu, CuETP1, Cu FRTP, Cu FRHC |
М1ф | Cu-DHP, SF-Cu |
М1P | Cu-DLP, SW-Cu |
М00к | OFC-Cu |
Главное свойство меди это-электропроводность, такие примеси как Висмут, Железо, Сурьма и Мышьяк ухудшают сопротивление Меди.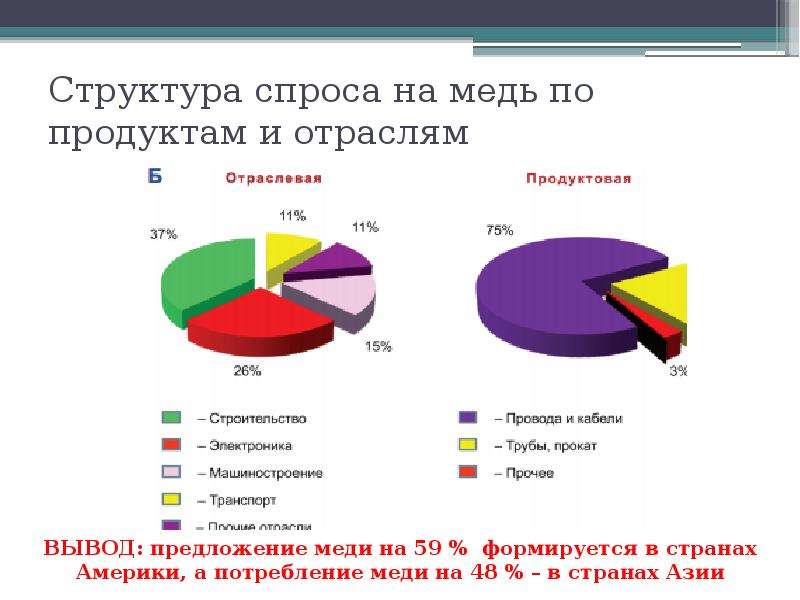 Таким образом из первой таблицы можно понять что для производства электротехнической меди необходимо гальваническое очищение исходного сырья
Таким образом из первой таблицы можно понять что для производства электротехнической меди необходимо гальваническое очищение исходного сырья
Для изготовления медной катанки при производстве КПП переплавляют медные катоды методом, который исключает насыщение меди кислородом. Медь переплавленная таким способом имеет марку М00 и М0. Стоимость электротехнических марок меди обусловлена высокими дополнительными затратами на ее рафинирование и очистку.
Катанка медная, бескислородная с высокой электропроводностью, светлая, не требующая дальнейшего травления, применяется для изготовления электротехнической проволоки.
Нормативный документ ТУ У 13614181.001-98, что соответствует ДСТУ EN 13601-СU-OF-R180-RND8.
Наименование | Значение |
Диаметр мм | 8,0 |
Отклонение по диаметру мм | +/- 0,2 |
Содержание Меди % | 99,98 |
Содержание кислорода % | 0,001 |
Удельное сопротивление при 20град. | 0,01724 |
Сопротивление разрыву Н/м.кв. | 160х108 |
Относительное удлинение % | 35 |
Число скручиваний-раскручиваний | 10 |
Число скручиваний до разрушения | 50 |
 омхмм.кв./м
омхмм.кв./м
Медь — Клиника 1
Медь – это жизненно важный микроэлемент, играющий существенную роль в синтезе гемоглобина и активации ферментов дыхательной цепи. Он входит в состав костей, хряща, соединительной ткани и миелиновых оболочек.
Основными источниками меди для человека являются такие продукты питания как орехи, шоколад, грибы, печень, злаки и сухофрукты.
Также медь может поступать в организм с водой, в случае, если она контактировала с медьсодержащими предметами (например с медьсодержащей посудой). После поступления в желудочно-кишечный тракт медь всасывается в тонкой кишке и соединяясь с белками крови, транспортируется в печень.
Недостаточность меди, как и ее избыток — редкие патологические состояния. Чаща встречается перенасыщение организма медью, связанное с нарушением ее обмена, либо хроническим отравлением.
Дефицит меди может внезапно возникнуть у людей, страдающих заболеваниями, вызывающими тяжелую мальабсорбцию (муковисцидозом, целиакией).
Недостаточное количество меди в крови грозит производством дефектных эритроцитов с низкой продолжительностью жизни, а также уменьшением активности ферментов, содержащих в своем составе этот микроэлемент.
Для чего используется исследование?
- Для диагностики болезни Вильсона-Коновалова (как правило совместно с тестом на церулоплазмин).
- Для оценки состояния пациента при подозрении на отравление медью, а также при ее недостатке или при нарушениях, влияющих на обмен меди (вместе с тестом на церулоплазмин).
- Для контроля за эффективностью лечения болезни Вильсона-Коновалова и патологического состояния, вызванного избытком меди или ее излишней потерей.


Когда назначается исследование?
Анализ назначается совместно с другими тестами (церулоплазмин), при наличии симптомов болезни Вильсона-Коновалова, подозрении на острое отравление медью, а также для оценки обеспеченности организма данным микроэлементом.
Стоимость исследования
Медь и её сплавы
Медь отличается высокими теплопроводностью, электропроводностью, коррозионной стойкостью, низкой температурой плавления, хорошо обрабатывается давлением, удовлетворительно резанием. Широко применятся в электротехнике, машино- и приборостроении. Медь по ГОСТ 859-78 выпускается в виде катодов, слитков, полос, лент, труб, проволоки, поковок, листов.
Латуни – двойные многокомпонентные медные сплавы с основным легирующим элементом – цинком. По сравнению с медью обладает более высокой прочностью и коррозионной стойкостью. Простые латуни обозначают буквой Л и цифрой, показывающей содержание меди в процентах. В специальных латунях после буквы Л пишут заглавную букву дополнительных легирующих элементов (А — алюминий, Б — бериллий, Ж — железо, К — кремний, Мц — марганец, Н — никель, О — олово, С — свинец, Ц — цинк, Ф. – фосфор) и через тире после содержания меди указывают содержание легирующих элементов в процентах. Латуни разделяют на литейные и деформируемые. Латуни, за исключением свинцовосодержащих, легко поддаются обработке давлением в холодном и горячем состоянии. Все латуни хорошо паяются твердыми и мягкими припоями.
Простые латуни обозначают буквой Л и цифрой, показывающей содержание меди в процентах. В специальных латунях после буквы Л пишут заглавную букву дополнительных легирующих элементов (А — алюминий, Б — бериллий, Ж — железо, К — кремний, Мц — марганец, Н — никель, О — олово, С — свинец, Ц — цинк, Ф. – фосфор) и через тире после содержания меди указывают содержание легирующих элементов в процентах. Латуни разделяют на литейные и деформируемые. Латуни, за исключением свинцовосодержащих, легко поддаются обработке давлением в холодном и горячем состоянии. Все латуни хорошо паяются твердыми и мягкими припоями.
Бронзами называют медные сплавы, в которых основными легирующими элементами являются различные металлы, кроме цинка. Маркируют бронзы буквами Бр, за которыми следуют заглавные буквы легирующих элементов, а через тире цифры, показывающие их процентное содержание.
По сравнению с латунью бронзы обладают более высокой прочностью, коррозионной стойкостью и антифракционными свойствами. Они весьма стойки на воздухе, в морской воде, растворах большинства органических кислот, углекислых растворах.
Они весьма стойки на воздухе, в морской воде, растворах большинства органических кислот, углекислых растворах.
Большинство бронз (за исключением алюминиевых) хорошо поддаются сварке и пайке твердыми и мягкими припоями.
См. также «Теоретический вес всех основных видов цветного металлопроката»
Медные сплавы.
Оловянные бронзы.
| Марка | Химический состав | Назначение | |||||
|---|---|---|---|---|---|---|---|
| Sn | P | Zn | Ni | Pb | |||
| обрабатываемые давлением (однофазные) по ГОСТ 5017–49 | |||||||
| Бр.ОФ6,5–0,15 | 6–7 | 0,1–0,25 | ― | ― | ― | Ленты, сетки в аппаратостроении, бумажнойпром. .Мембраны, пружины, детали работающие на трение. .Мембраны, пружины, детали работающие на трение. |
|
| Бр.ОЦ4–3 | 3,5 | ― | 2,7–3,3 | ― | ― | ||
| литейные (двухфазные) по ТУ | |||||||
| Бр.ОЦ10–2 | 9–11 | ― | 2–4 | ― | ― | шестерни, втулки, подшипники. | |
| Бр.ОФ10–1 | 9–11 | 0,8–0,12 | ― | ― | ― | То же, пластичность выше. | |
| Бр.ОНС11–4–3 | ― | ― | ― | 4 | 3 | То же, при нагреве. Втулки клапанов. | |
Алюминиевые бронзы (по ГОСТ 18175–72)
| Марка | Химический состав | Назначение | ||
|---|---|---|---|---|
| Al | Fe | Ni | ||
| высокой пластичности (однофазные) | ||||
Бр. А5 А5 |
4–6 | ― | ― | Ленты, полосы, для пружин. |
| высокой прочности (двухфазные) | ||||
| Бр.АЖ 9–4 | 8–10 | 2–4 | ― | Шестерни, втулки, арматура, в.т.ч для морской воды. |
| Бр.АЖН10–4–4 | 9,5–11 | 3,5–5,5 | 3,5–5,5 | То же, при больших давлениях и трении. |
Кремнистые бронзы (по ГОСТ 18175–72)
| Марка | Химический состав | Назначение | ||
|---|---|---|---|---|
| Si | Mn | Ni | ||
| Бр.КМц 3–1 | 2,75–3,5 | 1–1,5 | ― | Пружины, трубы, втулки в судостроении, авиации, химической промышленности. |
| Бр.КН 1–3 | 0,6–1,1 | 0,1–0,4 | 2,4–3,4 | Втулки, клапаны, болты,и др. детали для работы вморской и сточных водах. |
Бериллиевые бронзы (по ГОСТ 18175–72)
| Марка | Химический состав | Назначение | ||||
|---|---|---|---|---|---|---|
| Be | Ni | Ti | Mg | |||
| Бр.Б2 | 1,8–2,1 | 0,2–0,5 | ― | ― | Высокопрочные и токоведущие пружины, мембраны, сильфоны. | |
| Бр.БНТ1,7 | 1,6–1,85 | 0,2–0,4 | 0,1–0,25 | ― | ||
| Бр.БНТ1,9 | 1,85–2,1 | 0,2–0,4 | 0,1–0,25 | ― | ||
Бр. БНТ1,9Mr БНТ1,9Mr |
1,85–2,1 | 0,2–0,4 | 0,1–0,25 | 0,07–0,13 | ||
Латуни
| Марка | Химический состав | Назначение | ||||
|---|---|---|---|---|---|---|
| Cu | Al | Pb | Sn | другие | ||
| Простые латуни | ||||||
| Пластичные (однофазные), деформируемые в холодном и горячем состоянии | ||||||
| Л96 (томпак) | 95,0–97,0 | ― | ― | ― | ― | Трубки радиаторные, листы, ленты. |
| Л80 (полутомпак) | 79,0–81,0 | ― | ― | ― | ― | Трубки, лента, проволока. |
| Л68 | 67,0–70,0 | ― | ― | ― | ― | Листы, ленты для глубокой вытяжки. |
| Меньшей пластичности (двухфазные), деформируемые в горячем состоянии и литейные. | ||||||
| ЛС59–1 | 57,0–60,0 | ― | 0,8–1,9 | ― | ― | Листы, трубы, литье; хорошая обрабатываемость резанием. |
| Сложные латуни | ||||||
| Обрабатываемые давлением (однофазные) | ||||||
| ЛА 77–2 | 76,0–79,0 | 1,7–2,5 | ― | ― | ― | Трубы в морском и общем машиностроении |
| ЛО70–1 | 69,9–71,0 | ― | ― | 1–1,5 | ― | Трубы подгревателей |
| Литейные (двухфазные) по ГОСТ 17711–72 | ||||||
| ЛА 67–2,5 | 66–68 | 2–3 | <=1,0 | ― | ― | Отливки в морском и общем машиностроении |
| Сложные латуни повышенной прочности и стойкости против коррозии | ||||||
| ЛАН 59–3–2 | 57,0–60,0 | 2,5–3,5 | ― | ― | 2–3 Ni | Трубы, тяжело нагруженные детали в моторо- и судостроении |
| ЛАЖ 60–1–1 | 58,0–61,0 | 0,75–1,5 | <=0,4 | ― | 0,8–1,5 Fe | |
| Литейные (двухфазные) по ГОСТ 17711–72 | ||||||
| ЛМцЖ 55–3–1 | 53–58 | ― | <=0,5 | 1,3–4,5 | 0,5–1,5 Fe4–3 Mn | Массивное литье в судосроении.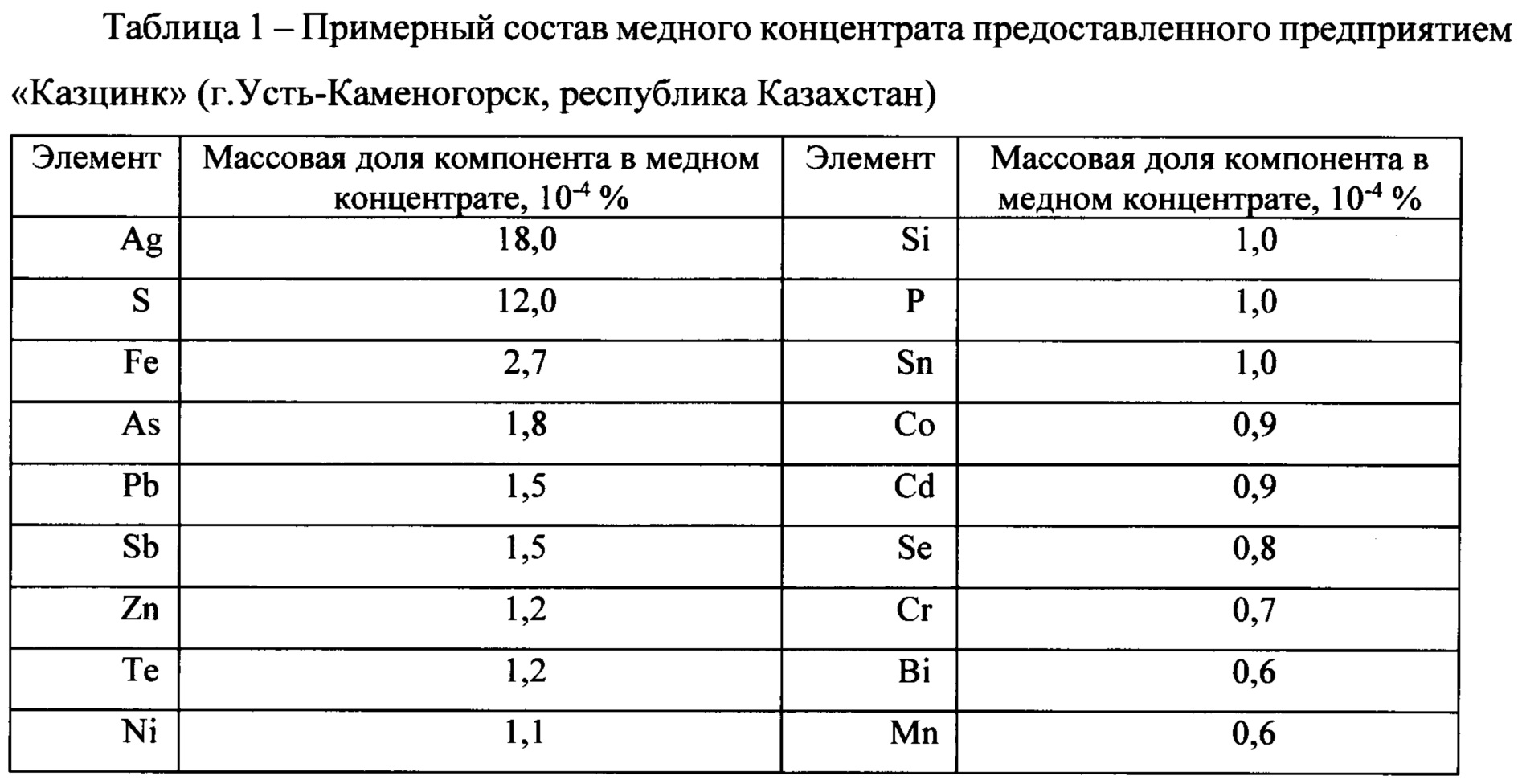 |
| ЛмцОС 58–2–2–2 | 57–60 | ― | 0,5–2,5 | 1,5–2,5 | 1,5–2,5 Mn | Шестерни, зубчатые колеса |
Концентрат медный
НАЗНАЧЕНИЕ И ОПИСАНИЕ
Концентрат медный получается при обогащении медьсодержащих руд и предназначен для производства металлической меди и извлечения сопутствующих ей полезных компонентов.
Концентрат медный (КМ) подразделяется на марки в зависимости от массовой доли меди. Химический состав марок КМ в пересчете на абсолютно сухое вещество должен соответствовать нормам, указанным в таблице.
ОСНОВНЫЕ ХАРАКТЕРИСТИКИ
| Марка | Содержание, % | ||
| Меди, не менее | Примесей, не более | ||
| цинка | свинца | ||
| КМ5 | 20 | 7 | 4,5 |
| КМ6 | 18 | 8 | 4,5 |
| КМ7 | 15 | 8,5 | 5 |
ПРИМЕЧАНИЕ
- По согласованию изготовителя с потребителем допускается в концентрате марки КМ7 массовая доля основного вещества не менее 12,0%;
- Массовая доля влаги в подсушенном медном концентрате не должна превышать 7,0%, в несушеном медном концентрате – не более 13,0%. Массовая доля влаги в медном концентрате, содержащим более 80% частиц класса менее 0,071мм или содержащем более 10% окислов меди, должна быть не более 14,0%;
- Массовая доля драгоценных металлов в медном концентрате определяют в каждой партии;
- Массовая доля редких металлов и серы определяется, если это предусмотрено договором на поставку, в котором указывают периодичность их определения.
 Массовая доля влаги в медном концентрате, содержащим более 80% частиц класса менее 0,071мм или содержащем более 10% окислов меди, должна быть не более 14,0%;
Массовая доля влаги в медном концентрате, содержащим более 80% частиц класса менее 0,071мм или содержащем более 10% окислов меди, должна быть не более 14,0%;УПАКОВКА И ТРАНСПОРТИРОВКА
Медный концентрат хранится насыпью в крытых складских помещениях.
Медный концентрат транспортируют железнодорожным транспортом насыпью в специализированных вагонах.
Возможна транспортировка водным и автомобильным видами транспорта в соответствии с правилами перевозок грузов, действующими на транспорте данного вида.
КАЧЕСТВО
ГОСТ Р 52998-2008 «Концентрат медный. Технические условия».
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности.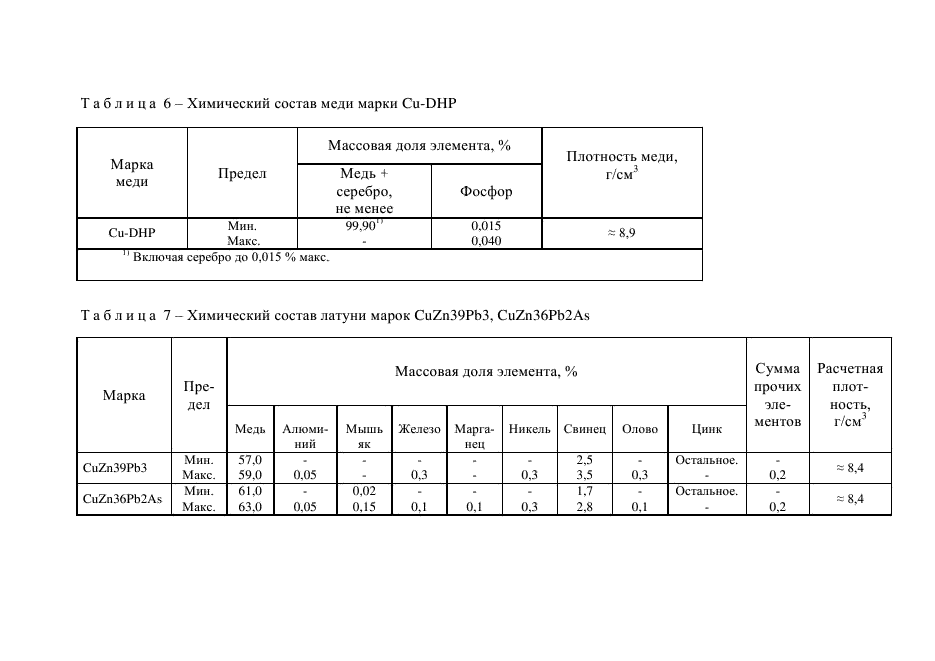 Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки вашего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.

Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки вашего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
 Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie
потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Состав медных сплавов, используемых греческой, этрусской и римской цивилизациями: 3. Происхождение и раннее использование латуни
Это третья из серии статей, посвященных составу медных сплавов, используемых в классическом мире.
Хотя известно несколько отдельных примеров доисторических медных сплавов, содержащих несколько процентов цинка, особенно на Кипре, сознательное производство латуни началось только в первом тысячелетии до нашей эры, вероятно, в Малой Азии. С 7 века греки говорили о латуни, но всегда как о дорогом и экзотическом металле, не производимом в Греции, и это подтверждается отсутствием цинка в подавляющем большинстве греческих бронз. Точно так же из многих проанализированных этрусских бронзовых изделий только две из 5-го и 3-го веков до нашей эры, соответственно, были из латуни, и они содержали около 11% цинка и менее 3% олова (см. Ниже).
С 7 века греки говорили о латуни, но всегда как о дорогом и экзотическом металле, не производимом в Греции, и это подтверждается отсутствием цинка в подавляющем большинстве греческих бронз. Точно так же из многих проанализированных этрусских бронзовых изделий только две из 5-го и 3-го веков до нашей эры, соответственно, были из латуни, и они содержали около 11% цинка и менее 3% олова (см. Ниже).
К I веку до нашей эры римляне использовали процесс цементации для производства латуни. Первоначально он, кажется, использовался для чеканки монет, но быстро стал популярным в других областях, особенно в декоративных металлических изделиях, где он в значительной степени заменил бронзу.В течение 2-го и 3-го веков нашей эры содержание цинка в монетах упало, и латунь с высоким содержанием цинка перестала использоваться, хотя латунь продолжала оставаться такой же популярной, как и когда-либо, составляя около 30% римских медных сплавов. К III веку сплав меди с содержанием свинца, цинка и олова по несколько процентов (современная «свинцовая бронза») использовался регулярно и остается таковым до сих пор.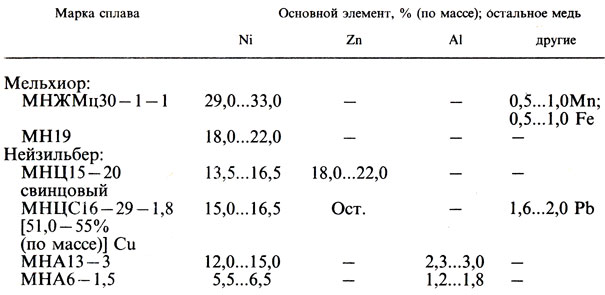
Со времен Библии короля Иакова 1611 года, в которой все медные сплавы называются «латунью», независимо от состава, раннее развитие и использование латуни было запутанным, и научные книги, как древние, так и современные, часто имели тенденцию к увеличению вместо того, чтобы уменьшить эту путаницу! Происхождение обоих терминов «латунь» и бронза неясно, но кажется, что в средневековые времена латунь применялась ко всем медным сплавам, и только с эпохи Возрождения термин bronzo использовался в Италии для обозначения медных сплавов, специально легированных оловом. .Однако эти термины продолжали неизбирательно использоваться в Англии до 19 века; Джонсон определяет бронзу как латунь в своем Словаре. Однако среди многих работ по этой теме работы Кейли (1964) и Вернера (1972) очень полезны и проливают свет на то, что в остальном является довольно темной областью.
Объединив информацию в этих книгах с новыми анализами, особенно анализами римского периода, которые будут опубликованы в последующих частях этой работы, мы получим гораздо более полную картину ранней истории меди с момента ее зарождения до конца римской эпохи. Империю можно построить.
Империю можно построить.
В данном обзоре под латунью понимается сплав меди, в который намеренно добавлен цинк. В доримской латуни цинк обычно заменяет олово, но в римской латуни оба металла часто присутствуют в сплаве.
Границы | Медь влияет на состав и функционирование микробных сообществ в морских биопленках в экологически значимых концентрациях
Введение
Загрязнение медью (Cu) в прибрежных районах в основном связано с бытовой и промышленной деятельностью (Parks et al., 2010; Oursel et al., 2013; Misson et al., 2016), а также использование противообрастающих красок на основе меди на корпусах судов (Yebra et al., 2004; Thomas and Brooks, 2010), особенно после запрета трибутилолова (TBT) в конце 1980-х годов во Франции. (Alzieu, 2000), а с 2003 года — в остальную Европу (Yebra et al., 2004). Повышенные уровни Cu можно обнаружить во многих частях мира, особенно возле закрытых гаваней и пристаней для яхт. Например, концентрации до 0,33 мкМ были обнаружены в заливе Сан-Диего, США (Schiff et al. , 2007) или до 0,41 мкМ в Тулонском заливе, Франция (Briand et al., 2017). На западном побережье Швеции уровни Cu были обнаружены при концентрациях до 5 мкг / л (0,08 мкМ) (Egardt et al., 2018), что превышает стандарт качества окружающей среды (EQS) для этого региона, то есть моря Каттегат. где был установлен EQS 4 мкг / л (0,07 мкМ) (HVMFS 2015: 4). Cu является важным элементом (Festa and Thiele, 2011), но становится токсичным при более высоких концентрациях (Amara et al., 2018), в зависимости от состава металла, накопления (Meylan et al., 2004; Serra et al., 2009) и подвергшегося воздействию организма (Barranguet et al., 2003; Amara et al., 2018). В фотосинтетически активных клетках Cu ингибирует фиксацию CO 2 и активность PSII (Pandey et al., 1992), вызывает окислительный стресс и в конечном итоге подавляет рост клеток (Gonçalves et al., 2018). У бактерий Cu влияет на различные клеточные ферменты и белки, участвующие в энергетическом обмене (Waldron et al., 2009). Cu влияет на видовой состав микробных сообществ, приводя к замене чувствительных таксонов на толерантные (Gustavson et al.
, 2007) или до 0,41 мкМ в Тулонском заливе, Франция (Briand et al., 2017). На западном побережье Швеции уровни Cu были обнаружены при концентрациях до 5 мкг / л (0,08 мкМ) (Egardt et al., 2018), что превышает стандарт качества окружающей среды (EQS) для этого региона, то есть моря Каттегат. где был установлен EQS 4 мкг / л (0,07 мкМ) (HVMFS 2015: 4). Cu является важным элементом (Festa and Thiele, 2011), но становится токсичным при более высоких концентрациях (Amara et al., 2018), в зависимости от состава металла, накопления (Meylan et al., 2004; Serra et al., 2009) и подвергшегося воздействию организма (Barranguet et al., 2003; Amara et al., 2018). В фотосинтетически активных клетках Cu ингибирует фиксацию CO 2 и активность PSII (Pandey et al., 1992), вызывает окислительный стресс и в конечном итоге подавляет рост клеток (Gonçalves et al., 2018). У бактерий Cu влияет на различные клеточные ферменты и белки, участвующие в энергетическом обмене (Waldron et al., 2009). Cu влияет на видовой состав микробных сообществ, приводя к замене чувствительных таксонов на толерантные (Gustavson et al. , 1999; Серра и др., 2009; Ancion et al., 2010). Подробные описания Cu-чувствительных и устойчивых к меди таксонов в экологических сообществах в настоящее время отсутствуют.
, 1999; Серра и др., 2009; Ancion et al., 2010). Подробные описания Cu-чувствительных и устойчивых к меди таксонов в экологических сообществах в настоящее время отсутствуют.
Перифитон, также называемый микрогитобентосом, образует биопленки самых разнообразных микробных сообществ, включая водоросли, бактерии, грибы и мейофауну, которые живут прикрепленными к подводным субстратам в водных экосистемах (Lock, 1993; Salta et al., 2013; Sanli et al. , 2015). Важность перифитона для водных экосистем связана с его функцией в качестве основного продуцента и его вкладом в биогеохимические циклы (Battin et al., 2003; Sundbäck et al., 2004). Использование перифитона для исследований в области экотоксикологии сообществ хорошо известно (Sabater et al., 2007; Eriksson et al., 2009a; Corcoll et al., 2015), поскольку он позволяет оценивать влияние загрязнителей на разных уровнях биологической организации. (Guasch et al., 2016). В этой связи подход, обычно используемый для обнаружения долгосрочных эффектов токсикантов в перифитонных сообществах, — это измерение PICT (терпимость сообщества, вызванного загрязнением), представленное Blanck et al.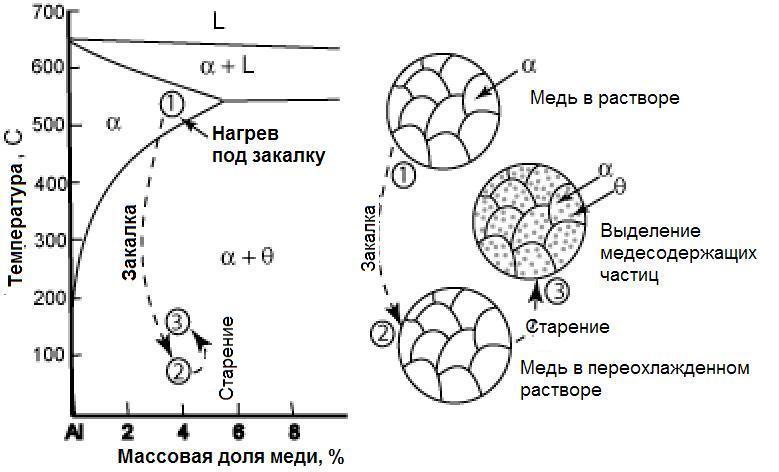 (1988). PICT основан на устранении микроорганизмов, чувствительных к рассматриваемому токсиканту, и индуцированной меж- и внутривидовой селекции организмов, которые более устойчивы к токсиканту. Все сообщество реструктурировано и, наконец, демонстрирует общее повышение устойчивости к токсиканту по сравнению с неэкспонированным эталонным сообществом. Эта индуцированная толерантность обычно количественно определяется как увеличение кратковременной ЕС50 всего сообщества по отношению к рассматриваемому токсиканту, что воспринимается как характеристика сообщества (Blanck et al., 1988; Corcoll et al., 2014; Тлили и др., 2015). Использование PICT для обнаружения воздействия Cu на морской и пресноводный перифитон оказалось более чувствительным, чем традиционные инструменты, основанные на составе сообщества, такие как наблюдения под микроскопом, подходы на основе профиля пигмента или отпечатки пальцев PCR-DGGE (Gustavson et al., 1999; Barranguet et al., 2003; Massieux et al., 2004; Serra et al., 2009; Tlili et al.
(1988). PICT основан на устранении микроорганизмов, чувствительных к рассматриваемому токсиканту, и индуцированной меж- и внутривидовой селекции организмов, которые более устойчивы к токсиканту. Все сообщество реструктурировано и, наконец, демонстрирует общее повышение устойчивости к токсиканту по сравнению с неэкспонированным эталонным сообществом. Эта индуцированная толерантность обычно количественно определяется как увеличение кратковременной ЕС50 всего сообщества по отношению к рассматриваемому токсиканту, что воспринимается как характеристика сообщества (Blanck et al., 1988; Corcoll et al., 2014; Тлили и др., 2015). Использование PICT для обнаружения воздействия Cu на морской и пресноводный перифитон оказалось более чувствительным, чем традиционные инструменты, основанные на составе сообщества, такие как наблюдения под микроскопом, подходы на основе профиля пигмента или отпечатки пальцев PCR-DGGE (Gustavson et al., 1999; Barranguet et al., 2003; Massieux et al., 2004; Serra et al., 2009; Tlili et al. , 2010).
, 2010).
Последние достижения в области секвенирования ДНК представляют собой мощный инструмент для обнаружения и количественной оценки воздействия токсичных веществ на экологические сообщества с высокой пропускной способностью выборки / наблюдения (Zhang et al., 2009; Ян и др., 2018). В частности, метабаркодирование ДНК, методика высокопроизводительного секвенирования ампликонов на основе ДНК, возникло как новый молекулярный инструмент для идентификации значительной части биологического сообщества, присутствующего в окружающей среде (Hebert et al., 2003). Новые вычислительные методы, применяемые к данным высокопроизводительного секвенирования ДНК, позволили разработать сетевой анализ совместной встречаемости для изучения потенциальных взаимодействий между таксонами (например, корреляции между таксонами). Эти новые аналитические методы позволили выйти за рамки классической ориентации на отдельные свойства микробных сообществ (например,g. состав и разнообразие сообщества, классически определяемые индексами α-разнообразия или ß-разнообразия; Barberán et al. , 2012). Например, в работе Мандаковича и др. По метабаркодированию. (2018) сетевой анализ применялся для выявления изменений в структуре и закономерностей совместной встречаемости в микробных сообществах почвы при различных стрессовых факторах окружающей среды.
, 2012). Например, в работе Мандаковича и др. По метабаркодированию. (2018) сетевой анализ применялся для выявления изменений в структуре и закономерностей совместной встречаемости в микробных сообществах почвы при различных стрессовых факторах окружающей среды.
Основная цель этого исследования — продемонстрировать полезность анализа ДНК-метабаркодирования для описания экотоксикологического эффекта Cu при концентрациях в окружающей среде на морские перифитные биопленки.Для этой цели использовалось метабаркодирование, чтобы описать влияние Cu на структуру сообщества прокариот и эукариот (т.е. биоразнообразие, состав сообщества, идентификацию чувствительных / толерантных таксонов и сеть сообществ). Кроме того, исследование направлено на изучение того, происходят ли вызванные Cu изменения в структуре сообщества одновременно с изменениями в функционировании сообщества (то есть фотосинтезом и толерантностью сообщества к Cu). Для этих целей природный морской перифитон подвергался воздействию меди в диапазоне концентраций (0. 01–10 мкМ) в полустатическом микромире в течение 18 дней. Мы выбрали гены 16S рРНК (область V3) и 18S рРНК (область V9) для нацеливания на прокариот и эукариот, соответственно. Наши результаты предоставляют новую информацию о том, как загрязнение Cu влияет на структуру и функционирование морских микробных сообществ, что может помочь в установлении соответствующих стандартов качества окружающей среды.
01–10 мкМ) в полустатическом микромире в течение 18 дней. Мы выбрали гены 16S рРНК (область V3) и 18S рРНК (область V9) для нацеливания на прокариот и эукариот, соответственно. Наши результаты предоставляют новую информацию о том, как загрязнение Cu влияет на структуру и функционирование морских микробных сообществ, что может помочь в установлении соответствующих стандартов качества окружающей среды.
Материалы и методы
Установка микромира и экспериментальный дизайн
Эксперимент проводился в помещении, в помещении с терморегулятором, на территории Центра морских наук Свена Ловена в Кристинеберге у фьорда Гульмар на западном побережье Швеции с 18 августа по 6 сентября 2015 года.Восемнадцать независимых микрокосмов, сделанных из прямоугольных стеклянных сосудов, вдохновленных тестом перифитона SWIFT, описанным Porsbring et al. (2007) были использованы для эксперимента. Каждый микрокосм содержал 300 мл природной морской воды, собранной из близлежащей нетронутой бухты (Госевикен: 58,245373 ° N, 11,433628 ° E). Морская вода с ее естественными организмами фильтровалась через сетку 200 мкм для удаления крупных организмов и обогащалась 0,7 мкМ фосфатом (KH 2 PO 4 ) и 0.8 мкМ нитрата (как NH 4 NO 3 ), чтобы избежать ограничения питательных веществ во время роста перифитона. Перифитону позволяли колонизировать прямоугольные слайды из полиэтилентерефталатгликоля (ПЭТГ) (6,9 × 1,4 см, 2 ). Каждый прямоугольный микромир имел стеклянный стержень, прикрепленный вдоль длинной стороны в середине дна сосуда, и 22 предметных стекла из ПЭТГ были помещены от нижнего стеклянного стержня к боковым стенкам сосуда, образуя угол ~ 22 ° между дном. и стенки сосуда.Морская вода покрывала половину поверхности горок. Вода из каждого микрокосма обновлялась каждый день. Чтобы стимулировать колонизацию и рост перифитона в начале эксперимента, инокулят морского перифитона готовили путем смахивания перифитона с верхней части 50–60 камней и гальки, собранных на максимальной глубине 60 см, в морскую воду.
Морская вода с ее естественными организмами фильтровалась через сетку 200 мкм для удаления крупных организмов и обогащалась 0,7 мкМ фосфатом (KH 2 PO 4 ) и 0.8 мкМ нитрата (как NH 4 NO 3 ), чтобы избежать ограничения питательных веществ во время роста перифитона. Перифитону позволяли колонизировать прямоугольные слайды из полиэтилентерефталатгликоля (ПЭТГ) (6,9 × 1,4 см, 2 ). Каждый прямоугольный микромир имел стеклянный стержень, прикрепленный вдоль длинной стороны в середине дна сосуда, и 22 предметных стекла из ПЭТГ были помещены от нижнего стеклянного стержня к боковым стенкам сосуда, образуя угол ~ 22 ° между дном. и стенки сосуда.Морская вода покрывала половину поверхности горок. Вода из каждого микрокосма обновлялась каждый день. Чтобы стимулировать колонизацию и рост перифитона в начале эксперимента, инокулят морского перифитона готовили путем смахивания перифитона с верхней части 50–60 камней и гальки, собранных на максимальной глубине 60 см, в морскую воду. Вода, камни и галька были взяты из той же нетронутой бухты, что и природная морская вода. Посевной материал энергично встряхивали и фильтровали через сито 200 мкм для удаления крупных организмов.Двадцать миллилитров инокулята с приблизительной концентрацией хлорофилла и 0,3 мкг мл -1 были введены дважды в каждый микрокосм в течение первой недели эксперимента. Микрокосмы инкубировали в терморегулируемой комнате при температуре 15 ° C. Суточный цикл свет / темнота продолжительностью 14/8 часов был смоделирован с помощью световых трубок OSRAM FLUORA с интенсивностью света на поверхности микрокосмов приблизительно 120 мкмоль фотонов m −2 с −1 .Микрокосмы находились в постоянном перемешивании с помощью горизонтальных встряхиваний.
Вода, камни и галька были взяты из той же нетронутой бухты, что и природная морская вода. Посевной материал энергично встряхивали и фильтровали через сито 200 мкм для удаления крупных организмов.Двадцать миллилитров инокулята с приблизительной концентрацией хлорофилла и 0,3 мкг мл -1 были введены дважды в каждый микрокосм в течение первой недели эксперимента. Микрокосмы инкубировали в терморегулируемой комнате при температуре 15 ° C. Суточный цикл свет / темнота продолжительностью 14/8 часов был смоделирован с помощью световых трубок OSRAM FLUORA с интенсивностью света на поверхности микрокосмов приблизительно 120 мкмоль фотонов m −2 с −1 .Микрокосмы находились в постоянном перемешивании с помощью горизонтальных встряхиваний.
План эксперимента включал неэкспонированные контрольные микрокосмы и 5 уровней воздействия Cu, каждый в трех экземплярах микромира. Номинальные уровни воздействия Cu были: 0,01, 0,06, 0,32, 1,78 и 10 мкМ. Исходные смеси Cu, в 1000 раз более концентрированные, чем номинальные, были приготовлены из реагента CuCl 2 · 2H 2 O (номер CAS: 10125-13-0, Sigma-Aldrich) с деионизированной водой. Триста микролитров исходных запасов CuCl 2 · 2H 2 O были добавлены к микрокосмам Cu, и такой же объем деионизированной воды был добавлен к неэкспонированным контролям, чтобы получить конечный объем 300 мл.Медь и деионизированная вода вводились одновременно с инокулятом перифитона для обработки Cu и контроля, соответственно. Температуру воды, pH, кислород и соленость периодически контролировали, по крайней мере, 10 раз в течение эксперимента, используя портативные мульти-зонды (HANNA Instruments). Вода всех микрокосмов отбиралась 3 раза до и после обновления воды для анализа на Cu. Для этого 50 мл воды фильтровали через 0,45 мкм, консервировали HNO 3 (65% выше чистой) до конечной концентрации 1% и выдерживали при 4 ° C до дальнейшего анализа с помощью ICP-MS.
Триста микролитров исходных запасов CuCl 2 · 2H 2 O были добавлены к микрокосмам Cu, и такой же объем деионизированной воды был добавлен к неэкспонированным контролям, чтобы получить конечный объем 300 мл.Медь и деионизированная вода вводились одновременно с инокулятом перифитона для обработки Cu и контроля, соответственно. Температуру воды, pH, кислород и соленость периодически контролировали, по крайней мере, 10 раз в течение эксперимента, используя портативные мульти-зонды (HANNA Instruments). Вода всех микрокосмов отбиралась 3 раза до и после обновления воды для анализа на Cu. Для этого 50 мл воды фильтровали через 0,45 мкм, консервировали HNO 3 (65% выше чистой) до конечной концентрации 1% и выдерживали при 4 ° C до дальнейшего анализа с помощью ICP-MS.
Отбор проб перифитона
Через 18 дней брали пробы перифитона для анализа концентрации хлорофилла и , фотосинтетических пигментов, фотосинтетической активности, устойчивости сообщества к Cu и микробного состава прокариот и эукариот. Для каждого микрокосма суспензия перифитона была получена путем соскабливания перифитона со слайдов в 150 мл морской воды, фильтрованной через 0,2 мкм и дополненной тем же количеством питательных веществ, что и в микрокосмах. Пять миллилитров суспензии перифитона фильтровали через фильтры Whatman GF / F и сразу использовали для анализов хлорофилла и .Десять миллилитров суспензии перифитона фильтровали через фильтры Whatman GF / F, замораживали при -20 ° C и хранили до экстракции пигментов. Десять миллилитров суспензии перифитона помещали в пробирки, осаждали центрифугированием при 6500 g в течение 10 мин при комнатной температуре, супернатант удаляли, а полученные осадки быстро замораживали в жидком азоте и хранили при -80 ° C до экстракции ДНК. Оставшуюся суспензию перифитона использовали для определения фотосинтетической активности ( 14 C-включение) и измерений толерантности в соответствии с подходом PICT.Анализы хлорофилла и , пигментных профилей и микробного состава были выполнены для всех обработок.
Для каждого микрокосма суспензия перифитона была получена путем соскабливания перифитона со слайдов в 150 мл морской воды, фильтрованной через 0,2 мкм и дополненной тем же количеством питательных веществ, что и в микрокосмах. Пять миллилитров суспензии перифитона фильтровали через фильтры Whatman GF / F и сразу использовали для анализов хлорофилла и .Десять миллилитров суспензии перифитона фильтровали через фильтры Whatman GF / F, замораживали при -20 ° C и хранили до экстракции пигментов. Десять миллилитров суспензии перифитона помещали в пробирки, осаждали центрифугированием при 6500 g в течение 10 мин при комнатной температуре, супернатант удаляли, а полученные осадки быстро замораживали в жидком азоте и хранили при -80 ° C до экстракции ДНК. Оставшуюся суспензию перифитона использовали для определения фотосинтетической активности ( 14 C-включение) и измерений толерантности в соответствии с подходом PICT.Анализы хлорофилла и , пигментных профилей и микробного состава были выполнены для всех обработок. По логистическим причинам измерения фотосинтеза и толерантности сообщества были выполнены только для контрольных микрокосмов и микрокосмов с экспозицией Cu 0,32 и 1,78 мкМ Cu.
По логистическим причинам измерения фотосинтеза и толерантности сообщества были выполнены только для контрольных микрокосмов и микрокосмов с экспозицией Cu 0,32 и 1,78 мкМ Cu.
Концентрация хлорофилла а и фотосинтетические пигменты
Хлорофилл a экстрагировали 10 мл этанола (96%) в течение 24 часов в темноте при комнатной температуре и количественно определяли флуорометрически (флуорометр Тернера 10-AU; конструкции Тернера, Саннивейл, Калифорния, США) по методам Джесперсена и Кристофферсен (1987).Фотосинтетические пигменты экстрагировали 4 мл смеси ацетон / метанол (80% / 20%, об. / Об.) При ультразвуковой обработке в ледяной суспензии в течение 3 мин. Девяносто микролитров экстрактов фильтровали через фильтры 0,45 мкм (фильтры VWR International Syringe) и анализировали с помощью высокоэффективной жидкостной хроматографии (HPLC; Shimadze Prominence HPLC Systems), следуя инструкциям Torstensson et al. (2015). Всего было идентифицировано 10 фотосинтетических пигментов, которые были выражены как отношение между площадью пика каждого идентифицированного пигмента и площадью пика хлорофилла и .Фукоксантин использовали в качестве пигментного маркера для группы водорослей Bacillarophyta, также известной как диатомовые водоросли (Roy et al., 2011).
Общественная терпимость, вызванная загрязнением (PICT)
Толерантность сообщества, вызванная загрязнением (PICT), была определена как увеличение значений EC50, определенных в тестах на краткосрочное ингибирование фотосинтеза с использованием 14 C-включения в качестве конечной точки согласно Eriksson et al. (2009b) с некоторыми модификациями. Из каждого исследуемого микрокосма были приготовлены три неэкспонированных контрольных образца путем смешивания 1 мл суспензии перифитона и 1 мл морской воды.На один образец из каждого микрокосма воздействовали 0,32, 1,35, 5,66, 23,8 и 100 мкМ Cu путем смешивания 1 мл суспензии перифитона с 1 мл растворов Cu. Морская вода, используемая для контроля, и растворы Cu фильтровали через 0,2 мкм и добавляли то же количество питательных веществ, что и в микрокосмах. Образцы смешивали в сцинтилляционных флаконах и инкубировали при 15 ° C и 120 мкмоль фотонов m -2 с -1 , осторожно встряхивая во время инкубации. Через 1 час к каждому образцу добавляли 50 мкл раствора бикарбоната натрия 14 C с радиоактивностью 80 мкКи / мл.Еще через час к каждому образцу добавляли 100 мкл формальдегида для прекращения фотосинтетической активности. Образцы подкисляли 100 мкл HCl, чтобы отогнать не включенный 14 ° C, и к каждому образцу добавляли 3 мл Instagel Plus. Дезинтеграции в минуту (DPM) рассчитывали из числа импульсов в минуту (CPM) на основе поправочных коэффициентов для характеристик закалки образца и эффективности машины с использованием жидкостного сцинтилляционного спектрометра (LS 500 Beckman). Абиотическое включение 14 C оценивали путем добавления 100 мкл формальдегида к одному образцу перед инкубацией с бикарбонатом натрия 14 C, и радиоактивность этого образца вычитали из радиоактивности других образцов из того же микромира. 14 С-включение использовали в качестве оценки фотосинтетической активности перифитона и выражали в относительных изменениях дезинтеграции в минуту (DPM). Толерантность сообщества была определена как разница в значениях EC 50 на основе фотосинтетической активности между контрольными микрокосмами, не подвергавшимися воздействию, и микрокосмами, подвергшимися воздействию 0,32 и 1,78 мкМ Cu.
Экстракция ДНК, ПЦР-амплификация и секвенирование
Микробный состав прокариот и эукариот оценивали с помощью метабаркодирования ДНК.Полную геномную ДНК экстрагировали с использованием набора Power Biofilm ® DNAIsolation Kit (MoBio Laboratories, США), следуя рекомендациям Corcoll et al. (2017). ДНК осаждали ацетатом натрия и этанолом перед последующим анализом. Бактериальные гены 16S рРНК (область V3) и гены эукариотической 18S рРНК (область V9) амплифицировали с использованием праймеров V3 (модифицированные праймеры 341F и 518R) (Klindworth et al., 2013) и праймеров V9 (1380F и 1510R) (Amaral-Zettler et al. al., 2009) соответственно. Для каждого образца проводили трехкратные реакции ПЦР, чтобы минимизировать потенциальную ошибку ПЦР.Библиотеки ампликонов ПЦР секвенировали с использованием технологии Ion Torrent Proton согласно протоколам производителя.
Биоинформатика
Для обработки необработанных последовательностей использовался конвейер QIIME v.1.8.0 (количественное понимание микробной экологии) (Caporaso et al., 2010). Последовательности низкого качества были обрезаны с помощью скрипта «split_libraries.py» с «-w 50 -q 20.» Химерная фильтрация ПЦР выполнялась с помощью скрипта «parallel_identify_chimeric_seqs.py» в QIIME с параметром по умолчанию.Оперативные таксономические единицы (OTU) были отобраны с отсечкой сходства последовательностей 97% после конвейера UPARSE (Edgar, 2013). Для каждой OTU была выбрана репрезентативная последовательность, а таксономия была назначена с использованием классификатора RDP (Wang et al., 2007) по базе данных Greengenes (DeSantis et al., 2006) и базы данных SILVA (Pruesse et al., 2007) для прокариот. и сообщество эукариотов соответственно. Пока нет единой таксономической иерархии эукариотических организмов (https: //www.arb-silva.де / проекты / эукариотическая таксономия /). В частности, некоторые таксоны не имеют четкой таксономической информации (например, класс, отряд, семейство и т. Д.). В некоторых группах есть промежуточные таксономические уровни, которых нет в других группах, таких как superphylum, subphylum или infraphylum. База данных содержит десятки тысяч таксонов, и очень сложно корректировать классификацию таксонов по одному. Следовательно, оригинальная классификация согласно SILVA была использована непосредственно в этом исследовании для эукаритоидных организмов. Необработанные данные были депонированы в архив краткого чтения NCBI (SRA) с идентификатором BioProject PRJNA496374, https: // www.ncbi.nlm.nih.gov/Traces/study/?acc=PRJNA496374.
Статистический анализ
Однофакторный дисперсионный анализ ANOVA использовался для оценки различий между обработками для концентрации хлорофилла и , фотосинтетической активности и значений EC 50 с использованием R (R Core Team, 2013). Эффективная концентрация Cu, которая имела 50% -ный эффект (EC 50 ), была определена после подгонки значений включения 14 C к модели доза-реакция (подгонка Weifull) с использованием пакета «drc» (Ritz et al., 2015) в R. Анализ главных компонентов (PCA) использовался для наблюдения за различиями в микробном составе прокариот и эукариот между обработками Cu с использованием взвешенных данных о расстоянии Unifrac в QIIME (Caporaso et al., 2010). Критерий Кайзера-Гуттмана использовался для определения значимости каждой оси анализа PCA. Поскольку матрица Unifrac включает филогенетическую информацию, PCA на основе расстояний Unifrac может лучше отражать различия в сообществах, чем матрица, основанная на данных о численности таксонов на уровне семейства или класса.Различия в составе прокариотического и эукариотического сообществ между обработками Cu оценивали с помощью MANOVA расстояний Unifrac. Различия между обработками Cu оценивали с помощью ANOVA с апостериорным тестом Даннета . Сети корреляции были созданы SparCC со 100 бутстрапами для присвоения двусторонних псевдо-значений p (Friedman and Alm, 2012). Для каждой обработки использовали 3 повтора для расчета корреляции Rho. Сети были отфильтрованы по величине корреляции> 0.6 и <-0,6, что указывает на сильные отношения совместной численности и совместного исключения. Сети визуализировались с помощью Cytoscape v 3.6.1, а топологические параметры сетей рассчитывались с помощью NetworkAnalyzer 2.7 (Assenov et al., 2008). Были получены следующие топологические параметры, узлы: количество сетей OTU, плотность сети: насколько плотно сеть заполнена ребрами (петли и дублированные ребра игнорируются), неоднородность сети: склонность сети содержать узловые узлы, централизация сети: распределение плотности сети, характерная длина пути: ожидаемое расстояние между двумя подключенными узлами, средн.количество соседей: среднее количество подключений узла в сети. Реакцию каждого таксона на воздействие Cu моделировали с помощью 3-параметрической лог-логистической модели и рассчитывали 50% -ную концентрацию эффектов (EC50). Бета-разнообразие оценивалось путем вычисления взвешенных расстояний UniFrac между образцами (Lozupone and Knight, 2007). Все образцы были разрежены на самой низкой глубине секвенирования, чтобы уменьшить систематические ошибки, возникающие из-за различий в глубине секвенирования (186400 и 112730 для сообществ эукариотов и прокариот, соответственно).
Результаты
Ответы перифитона после 18 дней воздействия пяти концентраций Cu от 0,01 до 10 мкМ в полустатическом тесте представлены ниже.
Условия эксперимента
Температура, соленость и pH были постоянными на протяжении всего эксперимента, изменяясь всего на 1–3% между ежедневными обновлениями воды. Средняя соленость составила 20,8 PSU, температура воды 17,9 ° C и pH 8,1 ( n = 59). Концентрации Cu в контроле и образцах с номинальной концентрацией 0.01 мкМ Cu были ниже предела количественного определения 0,02 мкМ. Для остальных образцов анализируемая Cu составляла от 33% до 90% номинальных концентраций, что было ближе к номинальным концентрациям при более высоких уровнях воздействия (дополнительная таблица 1). Поэтому номинальные концентрации используются для описания закономерностей реакции на концентрацию.
Воздействие на фотосинтетические пигменты, развитие толерантности населения и фотосинтетическую активность
В контрольных группах хлорофилл и составлял 52%, фукоксантин 12%, диадионоксантин + диатоксантин 11%, ß-каротены 9%, хлорофилл c 3%, зеаксантин 2% и остальные три не- идентифицированные пигменты составляли 11% от общего количества пигментов (среднее арифметическое из трех повторов) (рис. 1А).Концентрация хлорофилла и снижалась в зависимости от концентрации, достигая 81% ингибирования при максимальной концентрации 10 мкМ (рис. 1B). Аналогичная картина наблюдалась для общего содержания пигментов перифитона (рис. 1А). Напротив, относительное содержание фукоксантина увеличивалось в зависимости от концентрации, увеличивая до 60% от общего содержания пигмента при максимальном воздействии Cu, 10 мкМ Cu (рис. 1B). Фотосинтетическая активность во всех краткосрочных тестах обнаружения PICT подавлялась увеличением концентрации Cu, и значения EC50 можно было определить для всех микрокосмов, кроме одного из микрокосмов, подвергшихся воздействию 1.78 мкМ Cu. Краткосрочная EC50 (AVG ± SE) необработанного контроля составляла 8,88 ± 0,78 мкМ. После предварительного воздействия 0,32 мкМ Cu кратковременное значение EC50 увеличилось до 21,3 ± 1,34 мкМ, а после предварительного воздействия Cu при 1,78 мкМ оно увеличилось до 51,24 ± 5,85 мкМ (рис. 1B). То есть предварительное воздействие Cu увеличивало толерантность сообщества к краткосрочному воздействию на 2,3 и 5,7, соответственно, выше, чем при контрольной обработке, что указывает на повышение толерантности сообщества к воздействию меди. Фотосинтетическая активность (включение 14 C) при концентрациях 0.32 и 1,78 мкМ C было подавлено до 60% (рис. 1B).
Рисунок 1. (A) Профили пигментов (анализ ВЭЖХ), представленные как относительное содержание каждого пигмента (правая ось Y) и сумма всех пигментов (левая ось Y), (B) Хлорофилл a ( Концентрация Chl-a) (флуорометрический анализ) и фотосинтетическая активность в процентах от контроля показаны на левой оси Y, а толерантность сообщества к воздействию Cu показана на правой оси Y. Все значения представляют собой AVG ± SE; n = 3, кроме (B) , где EC50 для 1.78 было AVG ± SE; n = 2.
Влияние на состав прокариотических и эукариотических сообществ
ДНК-секвенирование фрагментов гена 16S и 18S дало в общей сложности 7 109 298 и 5 655 641 считывание высокого качества соответственно. Эти считывания сгруппированы в 17 445 прокариотических ОТЕ и 2151 эукариотических ОТЕ (Таблица 1). Как показывает индекс разнообразия Chao1 (Chao, 1984; дополнительный рисунок 1), глубина секвенирования была достаточной для достижения точки насыщения для идентификации таксонов прокариот и эукариот.
Таблица 1 . Информация о секвенировании для каждого образца.
В сообществах, не подвергавшихся воздействию, в сообществе прокариот преобладали классы Alphaproteobacteria и Flavobacteria, а также таксоны, связанные с Phycisphaerae и Saprosirae (рис. 2). В эукариотическом сообществе преобладали Ochrophyta и Metazoa, а также таксоны, связанные с Chlorophyta, Haptophyta и другими представителями Stramenopiles, чем Ochrophyta (рис. 2). Воздействие Cu уменьшило общее количество OTU (таблица 1), а также значительно уменьшило разнообразие хао1 как прокариотической, так и эукариотической части перифитона (дополнительный рисунок 1).Влияние Cu на состав прокариотического и эукариотического сообществ было подтверждено с помощью MANOVA на расстояниях Unifrac (Таблица 2). Существенные изменения в составе бактериального сообщества впервые наблюдались после воздействия 0,06 мкМ Cu, в то время как эукариотическая часть перифитона уже подверглась значительному воздействию после воздействия 0,01 мкМ Cu (таблица 2, рисунок 2). Эти результаты также отражены на графиках PCA, которые визуализируют влияние воздействия Cu на численность OTU (рис. 3). Для прокариот (рис. 3A) первая ось PCA объясняет 38% дисперсии и сгруппированы образцы из контроля и 0.01, 0,06, 0,32 и 1,78 мкМ Cu с правой стороны оси, тогда как образцы из 10 мкМ Cu были сгруппированы с левой стороны оси. Вторая ось PCA объяснила 37% дисперсии и в основном разделила образцы на три группы: (i) контрольные и низкие уровни воздействия Cu, (ii) 1,78 мкМ Cu и (iii) 10 мкМ Cu (рис. 3А). Третья ось также была значимой и составляла 12% без четкого разделения образцов. Примечательно, что при обработках, подвергшихся воздействию высоких уровней Cu (1.78 и 10 мкМ Cu), относительное количество прокариотических последовательностей, связанных с классами Nostocophycideae и Oscillatoriophycideae, было особенно высоким, а относительное количество последовательностей, связанных с Synechococcophycideae, было особенно низким (Рисунки 2A, 4).
Рисунок 2 . Гистограмма состава сообщества прокариотического перифитона на уровне класса (A) и эукариотического перифитона на уровне или более высоком таксономическом ранге (B) , полученная с помощью высокопроизводительного секвенирования на основе ампликонов.Обратите внимание, что обработка Cu: 0, 0,01, 0,06, 0,32, 1,78 и 10 в мкМ Cu. Stramenopiles_X: относится к OTU, связанным с другими членами Stramenopiles, кроме Ochrophyta.
Таблица 2 . Среднее одноразрывное расстояние между обработками Cu.
Рисунок 3 . Анализ главных компонентов (PCA) на основе взвешенных расстояний UniFrac для состава ОТЕ прокариотического сообщества (A) и эукариотического сообщества (B) в перифитоне.Обратите внимание, что обработка Cu: 0, 0,01, 0,06, 0,32, 1,78 и 10 в мкМ Cu.
Рисунок 4 . Обилие прокариот таксонов, связанных с геном 16S рРНК на разных таксономических уровнях (тип, класс и семейство), и корреляции Пирсона между концентрацией Cu и обилием таксонов. На рисунке показаны только таксоны, число прочтений которых превышает 2000. Обилие таксонов представлено на столбчатых диаграммах справа и включает все таксоны, обнаруженные в базе данных последовательностей.
Для Eukarytes первая ось PCA объясняет 68% дисперсии и разделяет образцы на основе возрастающего градиента воздействия Cu от левой стороны оси (контрольная обработка) до правого участка оси (максимальная обработка Cu , 10 мкМ Cu) (рис. 3В).Вторая ось объясняет примерно 12% дисперсии, а также разделяет образцы на основе возрастающего градиента воздействия Cu (рис. 3B). Следует отметить, что относительное количество последовательностей, ассоциированных с Ochrophyta и Lobosa, увеличивалось при обработке, подвергавшейся воздействию высоких уровней Cu (1,78 и 10 мкМ Cu; Рисунки 2A, 5). Напротив, относительное количество последовательностей, связанных с Haptophyta, Metazoa, Chlorophyt, Cliophora, Dinophyta и Stramenoplies, было заметно снижено по сравнению с контрольной обработкой (Рисунки 2A, 5).
Рисунок 5 . Обилие таксонов в эукариотах, связанных с геном 18S рРНК на разных таксономических уровнях (класс, тип или более высокая таксономическая группа), и корреляции Пирсона между концентрацией Cu и численностью таксонов. На рисунке показаны только таксоны, число прочтений которых превышает 2000.
Чувствительные и толерантные к Cu таксоны и изменения в общественной сети
Корреляционный анализ Пирсона показал различную чувствительность к Cu среди таксонов (рис. 4, 5).Как общая тенденция в таксонах прокариот (то есть на уровне типа), цианобактерии имели сильную положительную корреляцию с воздействием Cu, но, напротив, численность Planctomycetes и Proteobacteria phyla имела сильную отрицательную корреляцию с воздействием Cu (Рисунок 4). Для эукариот тип Stramenopiles_X сильно положительно коррелировал с воздействием Cu, а Amoebozoa слабо положительно коррелировал с воздействием Cu. Все другие типы внутри эукариот отрицательно коррелировали с воздействием Cu.В частности, Hacrobia и Alveolata показали сильную отрицательную корреляцию с воздействием Cu (Рисунок 5).
В общей сложности 23 таксона можно классифицировать как «чувствительные», демонстрируя явное уменьшение в зависимости от концентрации (дополнительный рисунок 2 и дополнительная таблица 2). Для прокариот наиболее чувствительными таксонами были филы Proteobacteria и Bacteriodetes. Большинство чувствительных таксонов имеют относительно высокие значения ЕС50 выше 1 мкМ Cu, за исключением четырех таксонов из Cytophagales, Rickettsiales, Myxococcales и Oceanospirillales, у которых значения EC50 ≤ 1 мкМ Cu (дополнительная таблица 2).Пять таксонов эукариот из таксономических групп Stramenopiles и Hacrobia были чувствительны к Cu со значениями EC50 ниже 2 мкМ Cu (дополнительная таблица 2).
Cu изменил сеть ассоциаций между таксонами в сообществах (рис. 6). В контроле и при обработке 0,01 мкМ Cu биовзаимодействия доминировали с Ciliophora, Dinophyta и Hilomonadea. Однако биовзаимодействия в основном осуществлялись цилиофорами в 1,78 и 10 мкМ Cu. Очень интересно, что при контроле и обработке с низким содержанием Cu виды взаимосвязаны, образуя замкнутый цикл.Когда концентрация Cu достигает 10 мкМ, взаимосвязь между видами становится открытой линейной. Низкое воздействие меди увеличивало сетевые ассоциации, но при концентрациях Cu 0,32 мкМ и выше количество узлов уменьшалось и было ниже, чем в контроле при 1,78 и 10 мкМ Cu (Таблица 3).
Рисунок 6 . Сеть между прокариотическими и эукариотическими сообществами при различных воздействиях Cu: (A) (контроль), (B) (0,01 мкМ Cu), (C) (0.06 мкМ Cu), (D) (0,32 мкМ Cu), (E) (1,78 мкМ Cu) и (F) (10 мкМ Cu). Связи между таксонами были получены с помощью корреляционного анализа «Пирсона». Зарезервированы только корреляции с корреляцией> 0,9 и «двусторонним» P <0,01. Коэффициенты корреляции между двумя узлами были помечены: положительный коэффициент - желтым, а отрицательный - черным. Размер узла указывает на «центральность промежуточности».
Таблица 3 .Топологические параметры сетевого анализа.
Обсуждение
Наше исследование микромира позволяет по-новому взглянуть на экологические последствия длительного воздействия Cu на морские прокариотические и микроэукариотические организмы в биопленках перифитона. Воздействие Cu длилось 18 суток, что намного больше времени генерации исследуемых микроорганизмов. Микробные прокариоты и эукариоты делятся во временном диапазоне от часов до нескольких дней. Следовательно, применяемое время воздействия считается для нашей тестовой системы долгосрочным или хроническим.Cu снижает прокариотическое и эукариотическое богатство (количество OTU) и количество их взаимодействий (количество узлов). Эукариоты оказались более чувствительными, чем таксоны прокариот. Несмотря на явные изменения в структуре сообществ, которые сделали подвергшиеся воздействию сообщества перифитона более устойчивыми к Cu, воздействие Cu снизило биомассу водорослей и фотосинтетическую активность экспонированных биопленок. Влияние на состав и функции сообщества наблюдалось при концентрации Cu 0,06 мкМ, которая, как известно, встречается в прибрежной среде Швеции (Egardt et al., 2018). Текущий стандарт качества окружающей среды для меди 0,07 мкМ (HVMFS 2015: 4), следовательно, не кажется достаточно защитным для перифитона, который содержит основных первичных продуцентов, особенно в прибрежных районах, где эвфотическая зона простирается до отложений (Wasmund, 1993; Sundbäck и др., 2004). Кроме того, следует подчеркнуть, что результаты были зарегистрированы в экологически реалистичных условиях, которые позволили экологической сукцессии и конкуренции сформировать сообщества в условиях длительного хронического воздействия Cu.Несмотря на то, что наше исследование проводилось в микрокосмах, микробный состав прокариот и эукариот, установленный в наших перифитных биопленках, был сопоставим с составом in situ морского перифитона на искусственных субстратах из того же региона (например, фьорд Гуллмар; Sanli et al., 2015; Corcoll et al., 2017).
Повышение устойчивости сообщества к загрязнению (PICT) совпало с изменениями в структуре и составе сообщества (рис. 1–3), а также со снижением биомассы водорослей и фотосинтетической активности (рис. 1).Это подтверждает точку зрения, впервые высказанную Blanck et al. (1988), толерантность сообщества возрастет, как только чувствительные виды и генотипы будут потеряны из сообщества. Эти наблюдения, однако, противоречат гипотезе функциональной избыточности, которая предполагает, что потеря видов мало влияет на экологические функции (Oliver et al., 2015). Вместо этого они поддерживают идею о том, что биоразнообразие должно быть полностью сохранено, чтобы гарантировать, что уязвимое сообщество может продолжать выполнять свои экологические функции в данной экосистеме (Tilman and Downing, 1994).Хотя предыдущие исследования биопленок установили связь между PICT и структурой микробных организмов (Dorigo et al., 2010) или генетическим составом фотосинтетических микроорганизмов (Eriksson et al., 2009a, b), это исследование направлено на то, чтобы насколько нам известно, первая статья, которая связывает PICT в биопленках с высокопроизводительными методами секвенирования ДНК (метабаркодирование), нацеленных на все прокариотические и микроэукариотические микроорганизмы.
Наблюдался широкий спектр чувствительных таксонов прокариот и эукариот (рис. 4, 5).Самая высокая толерантность к Cu среди прокариот была обнаружена у представителей типа Cyanobacteria, особенно у классов Nostocophycideae и Oscilaltoriphycideae (Рисунок 4). Устойчивость цианобактерий к воздействию Cu наблюдалась ранее (Barranguet et al., 2000; Serra et al., 2009) и может быть объяснена их способностью синтезировать внешние лиганды (Giner-Lamia et al., 2016), так что Cu накапливается внеклеточно (Serra et al., 2009).
Proteobacteria и Bacteroidetes phyla преобладали в неэкспонированном перифитоне, использованном в настоящем исследовании, который был взят из фьорда Гульмар на западном побережье Швеции.Это согласуется с предыдущими исследованиями морского бактериопланктона (Cottrell, Kirchman, 2000; Steven et al., 2012) и биопленок перифитона (Sanli et al., 2015; Corcoll et al., 2017). Толерантность Bacteroidetes к Cu наблюдалась в настоящем исследовании с биопленками, но аналогичные структуры наблюдались и у Bacteroidetes из отложений, подвергшихся воздействию Cu (Yang et al., 2018). Напротив, тип Proteobacteria был наиболее чувствительным к Cu. Численность 13 его таксонов, в основном из доминирующих классов альфа- и гамма-протеобактерий, была снижена в зависимости от концентрации, со значениями ЕС50 до 0.61 и 0,91 мкМ Cu соответственно (дополнительная таблица 2). Многие виды филума Proteobacteria ответственны за процессы нитрификации и денитрификации или связаны с ассимиляцией углерода (Ruiz-González et al., 2012; Sanli et al., 2015; Zhao et al., 2017). Учитывая высокую чувствительность Proteobacteria к Cu, мы поэтому предполагаем, что загрязнение Cu в морских районах может привести к нарушению круговорота азота. Следовательно, в следующем эксперименте было бы интересно изучить влияние Cu на микробную активность, связанную, например, с биогеохимическим циклом азота (Larsson et al., 2007; Mußmann et al., 2013), и не только по фотосинтезу.
Было обнаружено восемь различных эукариотических высших таксономических групп (Alveolata, Amoebozoa, Apusozoa, Archaeplastida, Hacrobia, Opisthokonta, Rhizaria и Stramenopiles; Рисунки 2, 5), что еще раз демонстрирует, что бентосные морские биопленки являются хозяевами высокого биоразнообразия прокариотических организмов ( Sanli et al., 2015; Corcoll et al., 2017). Stramenopiles и Amoebozoa были наиболее устойчивыми группами к воздействию Cu, хотя численность некоторых семейств грибов была в группе Opisthokonta (рис. 5).Bacillariophyta (диатомовые водоросли), семейство из группы Stramenopiles, было очень устойчиво к Cu (рис. 5). Пигментный анализ подтвердил эти результаты (рис. 1), поскольку относительное содержание фукоксантина, общего маркера диатомовых водорослей (Roy et al., 2011), также увеличивалось с увеличением концентрации Cu. Эти данные согласуются с предыдущими результатами (Gustavson et al., 1999), которые также сообщили об увеличении содержания центрических Bacillariophyta в морском фитопланктоне в результате воздействия Cu. Толерантность к Cu у диатомовых водорослей была связана с их способностью синтезировать внеклеточные полисахариды и фрустулин (Gonçalves et al., 2018). Тем не менее, ранее опубликованные исследования мезоксома также дают частично противоречивую картину толерантности диатомовых водорослей к Cu. Например, в исследованиях Barranguet et al. (2000) и Soldo and Behra (2000), диатомовые водоросли перифитона ручья были менее устойчивы к долгосрочному воздействию Cu, чем зеленые водоросли или цианобактерии. Различия, наблюдаемые между этими исследованиями, вероятно, объясняются тем, что в каждом исследовании микрокосма использовалось различное исходное сообщество водорослей. Первоначальный состав сообщества водорослей был описан как ключевой фактор развития устойчивости к металлам в сообществах водорослей (Pérez et al., 2010).
В грибах относительная численность большинства классов и семейств не зависела от воздействия Cu, что согласуется с предыдущими исследованиями мезокосмов отложений (Gardham et al., 2014; Yang et al., 2018). Описано несколько механизмов устойчивости грибов к токсичности Cu, такие как образование комплексов с медью компонентами клеточной стенки, изменения мембранного транспорта меди, синтез внутриклеточных медьсвязывающих металлотионеинов и фитохелатинов и образование внеклеточных комплексов с медью или — преципитирующие метаболиты (Cervantes and Gutierrezcorona, 1994).
Пять таксонов водорослей, где ингибирование зависело от концентрации: член семейства Pavlovaceae (Haptophyta), член отряда Erythropeltidales (Rhodophyta) и три таксона в пределах Stramenopiles, со значениями EC50 в диапазоне от 1,2 до 2 мкМ Cu (дополнительная таблица 2). Численность представителей Hacrobia сильно отрицательно коррелировала с воздействием меди (рис. 3, 5). Внутри Hacrobia относительная численность группы Haptophyta снизилась из-за воздействия Cu. Haptophyta — важная группа в океанах, особенно кальцифицирующие Haptophyta (кокколитофориды), которые оказывают сильное влияние на глобальные углеродные циклы (Tsuji and Yoshida, 2017).Обилие других групп водорослей (Chlorophyta, Rhodophyta и Dinophyta) также снизилось из-за воздействия Cu, но только в меньшей степени. Мы пришли к выводу, что воздействие Cu на вышеупомянутые классы водорослей вызвало наблюдаемое снижение общей биомассы водорослей и фотосинтетической активности (Рисунок 1), что хорошо согласуется с предыдущими исследованиями, которые продемонстрировали токсичность Cu для фотосинтеза и роста водорослей в низких концентрациях (например, Pérez и др., 2010).
Ciliphora, группа простейших в супертипе Alveolata, уменьшилась в численности под воздействием Cu (Рисунок 5), что подтверждает предыдущие результаты по чувствительности простейших к Cu (Madoni et al., 1996; Ян и др., 2018). Однако в целом о донных простейших известно немного. Использование метабаркодирования ДНК, по-видимому, способно преодолеть эти ограничения и предоставить новый инструмент для исследования простейших в экотоксикологических исследованиях.
Обилие большинства таксонов из таксономической группы Metazoa (Crustacea, Cnidaria или Rotifera) отрицательно коррелировало с воздействием Cu (Рисунок 5). Чувствительность Metazoa (т.е. нематод) к Cu и другим факторам окружающей среды хорошо известна (Bongers and Ferris, 1999; Boyd and Williams, 2003).Эта группа микроэукариот питается мелким органическим детритом, бактериями, водорослями, грибами и простейшими. Следовательно, они действуют как регуляторы разложения и, следовательно, играют ключевую роль в круговороте и динамике питательных веществ (Boyd and Williams, 2003; Stelzer, 2011). Следовательно, прямое воздействие Cu на Metazoa может влиять на трофические взаимодействия в биопленках перифитона, как показывает сетевой анализ (Рисунок 6). Относительная численность некоторых диатомовых водорослей (Bacillarophyta) в пределах типа Ochrophyta увеличивалась с воздействием Cu, как показывают данные по пигментам и метабаркодированию (Рисунки 2B, 3, 5).Тем не менее, Ochrophyta, как правило, исчезают из сетевого анализа — больше не взаимодействуя с узлами — при увеличении воздействия Cu (Рисунок 6, Таблица 3). Эти результаты предполагают, что взаимодействие бактерий и водорослей, которое, как известно, имеет важное значение, особенно в биопленках и микрофитобентосе (Decleyre et al., 2015; Krohn-Molt et al., 2017), отрицательно влияет на воздействие Cu. В целом, наш сетевой анализ показывает, что Cu изменила ассоциации между различными таксонами в прокариотических и эукариотических сообществах (Рисунок 6, Таблица 3), предполагая, что взаимодействия трофической цепи и микробная петля в биопленках перифитона будут изменены при определенных уровнях воздействия Cu.
Несмотря на то, что метабаркодирование ДНК стало популярным методом обнаружения большого количества таксонов в образцах окружающей среды (Hebert et al., 2003), этот метод также имеет свои ограничения. Выбор праймеров влияет на оценку биоразнообразия, и идеально универсальный праймер сложно или даже невозможно создать (Klindworth et al., 2013; Hugerth et al., 2014; Zhang et al., 2018). Чтобы преодолеть эти ограничения, комбинация многих специфических праймеров нацелена на каждое из царств эукариот (например,g., ген ITS для грибков; Nilsson et al., 2009, ген 23S, направленный на водоросли; Sherwood et al., 2008, или COI для беспозвоночных; Leray and Knowlton, 2015) может обеспечить улучшенное разрешение и меньшее смещение. В этом исследовании мы выбрали область V3 гена 16S рРНК для нацеливания на бактерии и область V9 18S рРНК для нацеливания на эукариот. Оба региона широко используются в метабаркодировании ДНК микробных сообществ в различных экосистемах (Amaral-Zettler et al., 2009; Klindworth et al., 2013; Corcoll et al., 2017; Ян и др., 2018). Однако область V4 гена 18S рРНК была предложена в качестве альтернативы области V9, чтобы захватить большее разнообразие (Pernice et al., 2013; Hugerth et al., 2014). Еще одним ограничением современных подходов к метабаркодированию, особенно в отношении морских микробных сообществ, является низкий охват общедоступных репозиториев последовательностей для многих естественных микроорганизмов, особенно микроэукариот (Bik et al., 2012; Sanli et al., 2015). Несмотря на неполноту справочных библиотек ДНК, недавно была продемонстрирована пригодность метабаркодирования для экологической оценки в пресноводных экосистемах с использованием бентических диатомовых водорослей (Rivera et al., 2018).
Таким образом, данное исследование позволило выявить изменения в составе сообществ бентосных про- и эукариот уже при 0,06 и 0,01 мкМ Cu соответственно. Эти концентрации воздействия являются реалистичными с экологической точки зрения (Egardt et al., 2018) и ниже действующих стандартов качества окружающей среды (EQS) для меди на западном побережье Швеции (HVMFS, 2015). Следовательно, наблюдаемые результаты в мезокосме предполагают, что текущие СКОС для морской среды не защищают прокариотических и эукариотических микробных организмов в морских биопленках.Наши результаты предоставляют новую информацию о том, как загрязнение Cu влияет на микробное биоразнообразие и состав сообществ в морской среде, данные, которые помогут установить соответствующие стандарты качества окружающей среды. Кроме того, эта работа демонстрирует надежность и многообещающий потенциал метабаркодирования ДНК как чувствительного инструмента для экотоксикологических исследований на уровне сообществ, который позволяет одновременно наблюдать воздействия на множество про- и эукариотических таксонов и, следовательно, выявлять особо чувствительные, непригодные для выращивания таксоны.
Авторские взносы
Разработку разработалиNC, KME и TB. NC и KME выполнили эксперимент, обработали образцы и провели анализ данных, не связанных с секвенированием. NC извлек ДНК. Секвенирование ампликона и биоинформатический анализ были выполнены JY и XZ. NC написал первый черновик статьи. Все авторы обсудили, интерпретировали результаты и внесли свой вклад в подготовку статьи.
Финансирование
Это исследование финансировалось Шведским исследовательским советом Formas (проект NICE, грант No.2011-1733, проект HerbEvol, грант № 2015–1464), Европейская комиссия (проект SOLUTIONS, соглашение о гранте 603437), шведский фонд Stiftelsen Brigit och Birger Wåhlströms minnesfond för den Bohuslänska havs- och insjömiljön и Шведская королевская академия наук ( КВА).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Авторы хотели бы поблагодарить всех сотрудников Центра морских наук им. Свена Ловена в Кристинеберге, а также студентов Хадриена Кроненбергера и Карину Платсен за их техническую помощь в проведении эксперимента.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fmicb.2018.03248/full#supplementary-material
Дополнительный рисунок S1 .Среднее ± стандартное отклонение (n = 3) индекса chao1 при каждой обработке медью (0, 0,01, 0,06, 0,32, 1,78 и 10 мкМ Cu) в зависимости от количества последовательностей, связанных с геном 16S рРНК (A) или с Ген 18S рРНК (B) .
Дополнительный рисунок S2 . Кривые зависимости реакции от концентрации для 23 таксонов, определенных как чувствительные к меди, их EC50, стандартное отклонение EC50. Назначение таксономии каждого таксона показано в дополнительной таблице 2.
Ссылки
Амара, И., Милед В., Слама Р. Б. и Ладхари Н. (2018). Противообрастающие процессы и токсическое воздействие необрастающих красок на морскую среду. Обзор. Environ. Toxicol. Pharmacol. 57, 115–130. DOI: 10.1016 / j.etap.2017.12.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Амарал-Цеттлер, Л. А., МакКлимент, Э. А., Даклоу, Х. У., и Хьюз, С. М. (2009). Метод изучения разнообразия протистанов с использованием массового параллельного секвенирования гипервариабельных участков V9 малых субъединичных генов рибосомной РНК. PLoS ONE 4: 06372. DOI: 10.1371 / journal.pone.0006372
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ансьон П. Ю., Лир Г. и Льюис Г. Д. (2010). Три основных металлических загрязнителя городских сточных вод (Zn, Cu и Pb) накапливаются в пресноводной биопленке и изменяют встроенные бактериальные сообщества. Environ. Загрязнение. 158, 2738–2745. DOI: 10.1016 / j.envpol.2010.04.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ассенов, Ю., Рамирес, Ф., Шелхорн, С. Э., Ленгауэр, Т., и Альбрехт, М. (2008). Вычисление топологических параметров биологических сетей. Биоинформатика 24, 282–284. DOI: 10.1093 / биоинформатика / btm554
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барберан А., Бейтс С. Т., Касамайор Э. О. и Фирер Н. (2012). Использование сетевого анализа для изучения моделей совместной встречаемости в микробных сообществах почвы. ISME J. 6, 343–351. DOI: 10.1038 / ismej.2011.119
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Барранге, К., Чарантони, Э., Планы, М., и Адмираал, В. (2000). Краткосрочная реакция моноспецифических и природных биопленок водорослей на воздействие меди. Eur. J. Phycol. 35, 397–406. DOI: 10.1080 / 09670260010001736001
CrossRef Полный текст | Google Scholar
Barranguet, C., van den Ende, F. P., Rutgers, M., Breure, A. M., Greijdanus, M., Sinke, J. J., et al. (2003). Медь-индуцированные модификации трофических отношений в биопленках речных водорослей и бактерий. Environ. Toxicol. Chem. 22, 1340–1349.
PubMed Аннотация | Google Scholar
Баттин, Т. Дж., Каплан, Л. А., Денис Ньюболд, Дж., И Хансен, К. М. Э. (2003). Вклад микробных биопленок в экосистемные процессы в мезокосмах ручьев. Природа 426, 439–442. DOI: 10.1038 / nature02152
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бик, Х. М., Поразинская, Д. Л., Крир, С., Капорасо, Дж. Г., Найт, Р., и Томас, В.К. (2012). Последовательность нашего пути к пониманию глобального биоразнообразия эукариот. Trends Ecol. Evol. 27, 233–243. DOI: 10.1016 / j.tree.2011.11.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Blanck, H., Wängberg, S. A., and Molander, S. (1988). «Толерантность сообщества, вызванная загрязнением — новый экотоксикологический инструмент», в Функциональное тестирование водной биоты для оценки опасности химических веществ , ред. Дж. Дж. Кэрнс и Дж. Р. Пратт (Филадельфия, Пенсильвания: Американское общество испытаний и материалов), 219–230.
Google Scholar
Бойд, В. А., и Уильямс, П. Л. (2003). Сравнение чувствительности трех видов нематод к меди и их использования в тестах на токсичность в воде и почве. Environ. Toxicol. Chem. 22, 2768–2774. DOI: 10.1897 / 02-573
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Briand, J. F., Barani, A., Garnier, C., Réhel, K., Urvois, F., LePoupon, C., et al. (2017). Пространственно-временные изменения сообществ морских биопленок, заселяющих искусственные субстраты, включая противообрастающие покрытия, в контрастных прибрежных средах Франции. Microb. Ecol. 74, 585–598. DOI: 10.1007 / s00248-017-0966-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010). соответствие QIIME позволяет анализировать данные секвенирования сообщества с высокой пропускной способностью. Nat. Методы 7, 335–336. DOI: 10.1038 / nmeth0510-335
CrossRef Полный текст | Google Scholar
Сервантес, К., и Гутьеррескорона, Ф.(1994). Механизмы резистентности к меди у бактерий и грибов. FEMS Microbiol. Ред. 14, 121–137.
PubMed Аннотация | Google Scholar
Чао, А. (1984). Непараметрическая оценка количества классов в генеральной совокупности. Сканд. J. Stat. 11: 265–270.
Google Scholar
Корколл, Н., Акуна, В., Барсело, Д., Каселлас, М., Гуаш, Х., Уэрта, Б. и др. (2014). Вызванная загрязнением толерантность сообщества к нестероидным противовоспалительным препаратам (НПВП) в сообществах речной биопленки, затронутых стоками очистных сооружений. Chemosphere 112, 185–193. DOI: 10.1016 / j.chemosphere.2014.03.128
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Корколл, Н., Каселлас, М., Уэрта, Б., Гуаш, Х., Акунья, В., Родригес-Мосас, С., и др. (2015). Влияние прерывистости потока и фармацевтического воздействия на структуру и метаболизм биопленок потока. Sci. Total Environ. 503–504. DOI: 10.1016 / j.scitotenv.2014.06.093
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Корколл, Н., Österlund, T., Sinclair, L., Eiler, A., Kristiansson, E., Backhaus, T., et al. (2017). Сравнение четырех методов экстракции ДНК для комплексной оценки бактериального разнообразия 16S рРНК в морских биопленках с использованием высокопроизводительного секвенирования. FEMS Microbiol. Lett. 364. DOI: 10.1093 / femsle / fnx139
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коттрелл, М. Т., и Кирчман, Д. Л. (2000). Состав сообщества морского бактериопланктона определяется с помощью библиотек клонов гена 16S рРНК и флуоресценции in situ гибридизации. Заявл. Environ. Microbiol. 66, 5116–5122. DOI: 10.1128 / aem.66.12.5116-5122.2000
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Decleyre, H., Heylen, K., Sabbe, K., Tytgat, B., Deforce, D., Van Nieuwerburgh, F., et al. (2015). Удвоение биомассы микрофитобентоса совпадает с десятикратным увеличением численности денитрификатора и общей численности бактерий в литоральных отложениях умеренного лимана. PLoS ONE 10: e0126583. DOI: 10,1371 / журнал.pone.0126583
PubMed Аннотация | CrossRef Полный текст | Google Scholar
ДеСантис, Т.З., Хугенгольц, П., Ларсен, Н., Рохас, М., Броди, Э.Л., Келлер, К. и др. (2006). Greengenes, проверенная химерами база данных генов 16S рРНК и рабочая среда, совместимая с ARB. Заявл. Environ. Microbiol. 72, 5069–5072. DOI: 10.1128 / AEM.03006-05
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дориго, У., Берард, А., Риме, Ф., Бушез, А., и Монтюэль, Б.(2010). In situ оценка восстановления перифитона в реке, загрязненной пестицидами. Aquat. Toxicol. 98, 396–406. DOI: 10.1016 / j.aquatox.2010.03.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эгардт, Дж., Мёрк Ларсен, М., Лассен, П., и Даллёф, И. (2018). Выбросы ПАУ и тяжелых металлов в прибрежную среду связаны с прогулочными судами. март Загрязнение. Бык. 127, 664–671. DOI: 10.1016 / j.marpolbul.2017.12.060
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эрикссон, К.М., Антонелли А., Нильссон Р. Х., Кларк А. К. и Бланк Х. (2009a). Филогенетический подход для обнаружения селекции на целевом участке противообрастающего соединения иргарола в толерантных сообществах перифитона. Environ. Microbiol. 11, 2065–2077. DOI: 10.1111 / j.1462-2920.2009.01928.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эрикссон, К. М., Кларк, А. К., Францен, Л. Г., Куйленшерна, М., Мартинес, К., и Бланк, Х. (2009b). Анализ на уровне сообществ последовательностей гена psbA и толерантности к иргаролу в морском перифитоне. Заявл. Environ. Microbiol. 75, 897–906. DOI: 10.1128 / AEM.01830-08
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гардам С., Хосе Г. К., Стефенсон С., Харитон А. А. (2014). Метабаркодирование ДНК соответствует экспериментальной экотоксикологии: расширение знаний об экологическом воздействии меди на пресноводные экосистемы, 1-е изд. Elsevier Ltd. doi: 10.1016 / B978-0-08-099970-8.00007-5
CrossRef Полный текст | Google Scholar
Гинер-Ламия, Дж., Перейра, С. Б., Бовеа-Марко, М., Футчик, М. Э., Таманьини, П., и Оливейра, П. (2016). Внеклеточные белки: новые ключевые компоненты устойчивости цианобактерий к металлам? Фронт. Microbiol. 7: 878. DOI: 10.3389 / fmicb.2016.00878
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гонсалвес, С., Калерт, М., Алмейда, С. Ф. П., и Фигейра, Э. (2018). Наука об общей окружающей среде, оценивающая воздействие КУ на пресноводные диатомеи: биохимические и метаболомные реакции tabellaria fl occulosa (Roth) Kützing. Sci. Total Environ. 625, 1234–1246. DOI: 10.1016 / j.scitotenv.2017.12.320
CrossRef Полный текст | Google Scholar
Guasch, H., Artigas, J., Bonet, B., Bonnineau, C., Canals, O., Corcoll, N., et al. (2016). «Использование биопленок для оценки воздействия химических веществ на пресноводные экосистемы», в Aquatic Biofilms: Ecology, Water Quality and Wastewater Treatment , ред. MD Romaní, AM Guasch и H. Balaguer (Норфолк: Caister Academic Press), 126 –144.
Google Scholar
Густавсон, К., Петерсен, С., Педерсен, Б., Стуэр-Лауридсен, Ф., Педерсен, С., и Венгберг, С.А. (1999). Толерантность сообщества, вызванная загрязнением (PICT), в сообществах прибрежного фитопланктона, подверженных воздействию меди. Hydrobiologia 416, 125–138. DOI: 10.1023 / A: 1003811419842
CrossRef Полный текст | Google Scholar
Hebert, P. D. N., Cywinska, A., Ball, S. L., and deWaard, J. R. (2003). Биологическая идентификация с помощью штрих-кодов ДНК. Proc. R. Soc. B Biol. Sci. 270, 313–321. DOI: 10.1098 / rspb.2002.2218
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Hugerth, L. W., Muller, E. E. L., Hu, Y. O. O., Lebrun, L. A. M., Roume, H., Lundin, D., et al. (2014). Систематический дизайн праймеров гена 18S рРНК для определения эукариотического разнообразия в микробных консорциумах. PLoS ONE 9: 095567. DOI: 10.1371 / journal.pone.0095567
PubMed Аннотация | CrossRef Полный текст | Google Scholar
HVMFS (2015).Havs- och vattenmyndighetens föreskrifterom ändring i Havs-och vattenmyndighetens föreskrifter (HVFS 2013: 19) по классификации и miljökvalitetsnormer avseende ytvatten. Hvmfs 19: 185.
Джесперсен, А.-М., и Кристофферсен, К. (1987). Измерения хлорофилла-а в фитопланктоне с использованием этанола в качестве экстракционного растворителя. Arch. Hydrobiol. 109, 445–454.
Google Scholar
Клиндворт, А., Прюсс, Э., Швир, Т., Пеплис, Дж., Кваст, К., Хорн М. и др. (2013). Оценка общих праймеров для ПЦР гена 16S рибосомной РНК для классических исследований и исследований разнообразия на основе секвенирования следующего поколения. Nucleic Acids Res. 41, 1–11. DOI: 10.1093 / nar / gks808
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Krohn-Molt, I., Alawi, M., Förstner, K.U., Wiegandt, A., Burkhardt, L., Indenbirken, D., et al. (2017). Понимание взаимодействий микроводорослей и бактерий в выбранных биопленках фикосферы с использованием метагеномного, транскриптомного и протеомного подходов. Фронт. Микробиол . 8: 1941. DOI: 10.3389 / fmicb.2017.01941
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ларссон, Ф., Петерсен, Д. Г., Даллоф, И., и Сундбэк, К. (2007). Комбинированное воздействие биоцида против обрастания и статуса питательных веществ на микробентосное сообщество мелководья. Aquat. Microb. Экол . 48, 277–294. DOI: 10.3354 / ame048277
CrossRef Полный текст | Google Scholar
Лерэ, М., Ноултон, Н. (2015).Штриховое кодирование ДНК и метабаркодирование стандартизованных образцов выявляют закономерности морского бентоса. Proc. Natl. Акад. Sci. США 112, 2076–2081. DOI: 10.1073 / pnas.1424997112
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лок, М. А. (1993). «Прикрепленные микробные сообщества в реках», в Aquatic Microbiology: An Ecological Approach , ed T. C. S. E. Fored (Oxford: Blackwell Scientific Publications), 113–138.
Мадони, П., Вескови, Л., и Эмилия, Р. (1996). Токсическое действие тяжелых металлов на сообщество простейших активного ила. Water Res. 30, 135–141.
Google Scholar
Мандакович Д., Рохас К., Мальдонадо Дж., Латорре М., Трэвисани Д., Делаж Е. и др. (2018). Структура и модели совместной встречаемости в микробных сообществах в условиях острого экологического стресса выявляют экологические факторы, способствующие устойчивости. Sci. Отчет 8: 5875. DOI: 10.1038 / s41598-018-23931-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Massieux, B., Boivin, M. E. Y., Ende, F. P., Van, D.en, Langenskio, J., and Marvan, P. (2004). Анализ структурных и физиологических профилей для оценки воздействия Cu на микробные сообщества биопленок. Заявл. Environ. Microbiol. 70, 4512–4521. DOI: 10.1128 / AEM.70.8.4512
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мейлан С., Бехра Р. и Сигг Л. (2004). Влияние видообразования металлов в естественной пресной воде на биоаккумуляцию меди и цинка в перифитоне: исследование микромира. Environ. Sci. Technol. 38, 3104–3111. DOI: 10.1021 / es034993n
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Misson, B., Garnier, C., Lauga, B., Dang, D.H., Ghiglione, J.F., Mullot, J.U., et al. (2016). Множественное химическое загрязнение способствует разнообразию бентосных прокариот в антропогенном Тулонском заливе. Sci. Total Environ. 556, 319–329. DOI: 10.1016 / j.scitotenv.2016.02.038
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мусманн, М., Рибо, М., фон Шиллер, Д., Мербт, С. Н., Аугспургер, К., Карваутц, К. и др. (2013). Колонизация пресноводных биопленок нитрифицирующими бактериями из активного ила. FEMS Microbiol. Экол . 85, 104–115. DOI: 10.1111 / 1574-6941.12103
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нильссон, Р. Х., Риберг, М., Абаренков, К., Шёквист, Э., и Кристианссон, Э. (2009). Регион ITS как объект для характеристики грибных сообществ с использованием новых технологий секвенирования. FEMS Microbiol. Lett. 296, 97–101. DOI: 10.1111 / j.1574-6968.2009.01618.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Оливер Т. Х., Херд М. С., Исаак Н. Дж. Б., Рой Д. Б., Проктер Д., Эйгенброд Ф. и др. (2015). Биоразнообразие и устойчивость функций экосистем. Trends Ecol. Evol. 30, 673–684. DOI: 10.1016 / j.tree.2015.08.009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Урсель, Б., Гарнье, К., Дурье, Г., Мунье, С., Оманови, Ч, Д., и Лукас, Ю. (2013). Динамика и судьба микроэлементов металлов, хронически поступающих в прибрежную зону Средиземного моря, подверженную влиянию крупных городских территорий. март Загрязнение. Бык. 69, 137–149. DOI: 10.1016 / j.marpolbul.2013.01.023
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Панди П. К., Сингх С. Б. и Сингх С. П. (1992). Поглощение Cu цианобактериями: судьба отдельных фотохимических реакций. Curr. Microbiol. 24, 35–39.DOI: 10.1007 / BF01570097
CrossRef Полный текст | Google Scholar
Паркс Р., Донье-Марешаль М., Фрикерс П. Э., Тернер А. и Ридман Дж. У. (2010). Биоциды против обрастания в отброшенных частицах морской краски. март Загрязнение. Бык. 60, 1226–1230. DOI: 10.1016 / j.marpolbul.2010.03.022.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перес П., Бейрас Р. и Фернандес Э. (2010). Мониторинг токсичности меди в сообществах естественного фитопланктона: применение быстрой флуорометрии. Ecotoxicol. Environ. Saf. 73, 1292–1303. DOI: 10.1016 / j.ecoenv.2010.06.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пернис М.С., Логарес Р., Гийу Л. и Массана Р. (2013). Общие закономерности разнообразия основных линий морских микроэукариот. PLoS ONE 8: e57170. DOI: 10.1371 / journal.pone.0057170
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Порсбринг, Т., Аррениус, А., Бакхаус, Т., Kuylenstierna, M., Scholze, M., and Blanck, H. (2007). Тест SWIFT на перифитон для высокопроизводительной оценки токсического воздействия на развитие сообщества микроводорослей. J. Exp. Mar. Biol. Ecol. 349, 299–312. DOI: 10.1016 / j.jembe.2007.05.020
CrossRef Полный текст | Google Scholar
Pruesse, E., Quast, C., Knittel, K., Fuchs, B.M., Ludwig, W., Peplies, J., et al. (2007). SILVA: всеобъемлющий онлайн-ресурс для проверенных и согласованных данных о последовательностях рибосомных РНК, совместимых с ARB. Nucleic. Acids Res. 35, 7188–7196. DOI: 10.1093 / nar / gkm.864
PubMed Аннотация | CrossRef Полный текст | Google Scholar
R Основная команда (2013). R: Язык и среда для статистических вычислений . Вена: Фонд R для статистических вычислений. Доступно в Интернете по адресу: http // www.R-project.org /
Ривера, С. Ф., Васселон, В., Жаке, С., Бушез, А., Аризтеги, Д., и Риме, Ф. (2018). Метабаркодирование озерных бентосных диатомовых водорослей: от структурных сообществ до экологической оценки. Hydrobiologia 807, 37–51. DOI: 10.1007 / s10750-017-3381-2
CrossRef Полный текст | Google Scholar
Рой С., Ллевеллин К., Эгеланн Э. и Йонсен Г. (ред.). (2011). Пигменты фитопланктона: характеристика, хемотаксономия и применение в океанографии (Кембриджская серия по химии окружающей среды). Кембридж: Издательство Кембриджского университета. DOI: 10.1017 / CBO9780511732263
CrossRef Полный текст | Google Scholar
Руис-Гонсалес, К., Лефорт, Т., Массана, Р., Сим, О, Р., и Газоль, Дж. М. (2012). Diel изменения основной массы и гетеротрофной активности одноклеточных бактерий в зимних поверхностных водах северо-западной части Средиземного моря. Лимнол. Oceanogr. 57, 29–42. DOI: 10.4319 / lo.2012.57.1.0029
CrossRef Полный текст | Google Scholar
Sabater, S., Guasch, H., Ricart, M., Romaní, A., Vidal, G., Klünder, C., et al. (2007). Мониторинг воздействия химикатов на биологические сообщества. Биопленка как интерфейс. Анал. Биоанал. Chem. 387, 1425–34. DOI: 10.1007 / s00216-006-1051-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Salta, M., Wharton, J. A., Blache, Y., Stokes, K. R., and Briand, J.-F. (2013). Морские биопленки на искусственных поверхностях: структура и динамика. Environ. Microbiol. 15, 2879–2893. DOI: 10.1111 / 1462-2920.12186
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Санли, К., Бенгтссон-Пальме, Дж., Нильссон, Р. Х., Кристианссон, Э., Альм Розенблад, М., Бланк, Х. и др. (2015). Метагеномное секвенирование морского перифитона: таксономическое и функциональное понимание сообществ биопленок. Фронт. Microbiol. 6: 01192. DOI: 10.3389 / fmicb.2015.01192
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шифф К., Браун Дж., Диль Д. и Гринштейн Д. (2007). Степень и масштабы загрязнения медью в маринах в районе Сан-Диего, Калифорния, США. мар.Загрязнение. Бык. 54, 322–328. DOI: 10.1016 / j.marpolbul.2006.10.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Серра А., Корколл Н. и Гуаш Х. (2009). Накопление меди и токсичность в речном перифитоне: влияние истории воздействия. Chemosphere 74, 633–641. DOI: 10.1016 / j.chemosphere.2008.10.036
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шервуд А. Р., Чан Ю. Л. и Престинг Г.Г. (2008). Применение универсально амплифицирующих пластидных праймеров для отбора проб из окружающей среды сообщества перифитона ручья. Mol. Ecol. Ресурс. 8, 1011–1014. DOI: 10.1111 / j.1755-0998.2008.02138.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сольдо Д. и Бехра Р. (2000). Долговременное влияние меди на структуру сообществ перифитона пресной воды и устойчивость к меди, цинку, никелю и серебру. Aquat. Toxicol. 47, 181–189.DOI: 10.1016 / S0166-445X (99) 00020-X
CrossRef Полный текст | Google Scholar
Стельцер, К. П. (2011). Первая оценка разнообразия размеров генома у коловраток-моногононов. Hydrobiologia 662, 77–82. DOI: 10.1007 / s10750-010-0487-1
CrossRef Полный текст | Google Scholar
Стивен Б., Макканн С. и Уорд Н. Л. (2012). Пиросеквенирование генов пластидной 23S рРНК выявило разнообразные и динамичные популяции цианобактерий и водорослей в двух эвтрофных озерах. FEMS Microbiol. Ecol. 82, 607–615. DOI: 10.1111 / j.1574-6941.2012.01429.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сундбэк, К., Линарес, Ф., Ларсон, Ф., Вульф, А., и Энгельсен, А. (2004). Бентические потоки азота вдоль градиента глубины в микропливном фьорде: роль денитрификации и Микрофитобентос 49, 1095–1107. DOI: 10.4319 / lo.2004.49.4.1095
CrossRef Полный текст | Google Scholar
Тильман, Д.и Даунинг Дж. А. (1994). Биоразнообразие и стабильность пастбищ. Природа 367, 363–365. DOI: 10.1038 / 367363a0
CrossRef Полный текст | Google Scholar
Тлили А., Берард А., Бланк Х., Бушез А., Кассио Ф., Эрикссон К. М. и др. (2015). Общественная толерантность, вызванная загрязнением (PICT): к экологически значимой оценке риска химических веществ в водных системах. Freshw. Биол. 61, 2141–2151. DOI: 10.1111 / fwb.12558
CrossRef Полный текст | Google Scholar
Тлили, А., Берар, А., Рулье, Ж.-Л., Вола, Б., и Монтюэль, Б. (2010). PO43 — зависимость толерантности автотрофных и гетеротрофных сообществ биопленок к меди и диурону. Aquat. Toxicol. 98, 165–77. DOI: 10.1016 / j.aquatox.2010.02.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Торстенссон, А., Динаске, Дж., Кьеричи, М., Франссон, А., Риман, Л., и Вульф, А. (2015). Физико-химический контроль состава и разнообразия сообществ бактерий и простейших в морском льду Антарктики. Environ. Microbiol. 17, 3869–3881. DOI: 10.1111 / 1462-2920.12865
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цудзи Ю. и Ёсида М. (2017). Биология гаптофитов: сложные клеточные процессы, управляющие глобальным углеродным циклом. Adv. Бот. Res. 84, 219–261. DOI: 10.1016 / bs.abr.2017.07.002
CrossRef Полный текст | Google Scholar
Ван, К., Гаррити, Г. М., Тидже, Дж. М., и Коул, Дж. Р. (2007). Наивный байесовский классификатор для быстрого отнесения последовательностей рРНК к новой бактериальной таксономии. Заявл. Environ. Microbiol. 73, 5261–5267. DOI: 10.1128 / AEM.00062-07
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Васмунд, Н. (1993). Экология и биопродукция в микрофитобентосе цепи мелких заливов (Бодденс) к югу от полуострова Дарсс-Цингст (южная часть Балтийского моря). Внутр. Rev. Total Hydrbiol. 71, 153–178. DOI: 10.1117 / 12.813996
CrossRef Полный текст | Google Scholar
Янг Дж., Се Й., Йеппе К., Лонг, С., Петтигров, В., Чжан, X. (2018). Чувствительные реакции микробиоты на медь в тесте на токсичность донных отложений. Environ. Toxicol. Chem. 37, 599–608. DOI: 10.1002 / etc.3980
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Йебра, Д. М., Киил, С., Дам-Йохансен, К. (2004). Технология против обрастания — Прошлые, настоящие и будущие шаги в направлении создания эффективных и экологически чистых противообрастающих покрытий. Prog. Орг. Покрытия 50, 75–104.DOI: 10.1016 / j.porgcoat.2003.06.001
CrossRef Полный текст | Google Scholar
Чжан, X., Ся, П., Ван, П., Ян, Дж., И Бэрд, Д. Дж. (2018). Омикс прогрессирует в экотоксикологии. Environ. Sci. Technol. 52, 3842–3851. DOI: 10.1021 / acs.est.7b06494
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжао, Дж., Ву, Дж., Ли, X., Ван, С., Ху, Б., и Дин, X. (2017). Характеристики денитрификации и микробное сообщество в катоде мтц с аэробной денитрификацией при высоких температурах. Фронт. Microbiol. 8: 9. DOI: 10.3389 / fmicb.2017.00009
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Стратегия проектирования с ориентацией на свойства для высокопроизводительных медных сплавов с помощью машинного обучения
Построение и анализ двух моделей BP NN
Первая модель BP NN (C2P) обучена предсказывать свойства UTS и EC на основе составов медных сплавов на условие сохранения обработки без изменений. На входе — составы, а на выходе — UTS и EC, которые показаны на рис.S1 и рис. S2. Более подробная информация о модели C2P представлена в дополнительной информации.
На рис. 1a – d представлены графики прогнозируемых значений UTS и EC из модели в зависимости от измеренных значений UTS и EC для обучающего набора, набора проверки, набора тестов и всех данных, соответственно. Точки данных располагаются вдоль диагональной линии (как показано пунктирной линией на каждой панели), указывая на то, что прогнозы вполне согласуются с измеренными значениями. Далее мы линейно подогнали прогнозируемые значения к измеренным значениям, как показано сплошной линией.Чем ближе сплошная линия к диагональной пунктирной линии, тем лучше производительность модели. Видно, что наша модель очень хорошо работает с отдельными наборами данных и со всеми данными.
Рис. 1Прогнозируемые значения UTS и EC из модели состав → свойство (C2P) как функция измеренных значений UTS и EC для a обучающего набора, b набора проверки, c набор тестов, и d все данные. (Пунктирная линия представляет выходные данные модели точно так же, как целевые данные в наборе данных; сплошная линия представляет результаты регрессии между выходными данными и целевыми данными.)
Обратный расчет состава, обработки и микроструктуры сплавов с заданными свойствами значительно сократит время и затраты на разработку новых материалов и является конечной целью материаловедения. 25 Самым большим препятствием является сложная взаимосвязь между композицией, обработкой, структурой и производительностью. Когда условия обработки задаются постоянно, состав является доминирующей переменной, определяющей микроструктуру и свойства сплава.Поэтому мы строим модель BP NN (P2C) с входом UTS и EC и выходом легирующих элементов, кроме Cu, которые были показаны на рис. S3 и рис. S4. Более подробная информация о модели P2C представлена в дополнительной информации.
Диаграммы разброса на рис. 2 показывают, что прогнозируемые значения содержания очень согласуются с измеренными значениями и падают вдоль пунктирных диагональных линий. Более того, линейно аппроксимируемые сплошные кривые очень близки к пунктирным диагональным линиям. Таким образом, P2C BP NN работает достаточно хорошо и может использоваться для рекомендации новых композиций.Модель P2C может одновременно выводить набор из 10 значений концентрации элементов во время работы. Например, когда мы вводим значения UTS и EC в модель P2C, модель P2C выводит значения концентрации 10 элементов одновременно. Набор из 10 значений концентрации элементов называется схемой расчета состава сплава.
Рис. 2Предсказанные значения содержимого из модели свойство → композиция (P2C) как функция измеренных значений содержимого для a обучающего набора, b набора проверки, c набора тестов, и d все данные.(Пунктирная линия представляет выходные данные модели точно так же, как целевые данные в наборе данных; сплошная линия представляет результаты регрессии между выходными данными и целевыми данными.)
Мы дополнительно проверяем точность прогноза Модель C2P и модель P2C путем сравнения прогнозов с экспериментальными данными, которых нет в нашей базе данных. Производительность на обучающих данных также включена в этот раздел. В таблице S1 дополнительной информации перечислены среднеквадратичная ошибка (RMSE) модели C2P и модели P2C, обученной с одной и той же базой данных, а также данные корреляции регрессионного соответствия (R) между прогнозируемой производительностью и целевой производительностью при различных наборах данных.Точность ошибки обучения сети C2P и коэффициенты корреляции подгонки регрессии на проверочном наборе и наборе тестов выше, чем в модели P2C. Приведенные выше результаты показывают, что обе модели хорошо работают с точки зрения производительности обучения, а модель C2P по-прежнему лучше, чем модель P2C. Основные причины этого различия обсуждаются ниже.
Чтобы проверить прогностическую эффективность модели C2P, мы выбрали сплавы, которые не представлены в нашей базе данных. На свойства медных сплавов влияет множество факторов, особенно на условия обработки.Сплав с одинаковым составом может иметь разную микроструктуру, полученную при разных условиях обработки, что приводит к большим отклонениям в свойствах. Мы используем лучшие свойства этих сплавов с таким же составом после обработки на твердый раствор, деформации и старения для проверки нашей модели C2P.
Прогнозируемые и экспериментальные значения для этих сплавов сравниваются в таблице 1. Сплав Cu – 2,93Ni – 0,9Si – 0,13Mg – 0,53Zn является коммерчески выпускаемым продуктом, а Cu – 2,20Ni – 0,42Si – 0.Сплав 08Mg – 0.30Zn производится и измеряется в нашей собственной лаборатории, а остальные четыре сплава 26,27,28 были опубликованы совсем недавно. Эти данные не включены в нашу базу данных. Таблица 1 показывает, что отклонение между прогнозируемыми и экспериментальными значениями находится в пределах 5,0%, что указывает на то, что модель C2P может точно прогнозировать свойства сплавов.
Таблица 1 Проверка прогноза характеристик сплава на основе состава (модель C2P)Для проверки точности состава сплава, разработанного моделью P2C, мы установили целевое значение UTS как 500, 550, 600, 650 и 700. МПа, соответственно, вместе с целевым значением ЕС 50.0% МАКО. Эти свойства вводятся в модель P2C, и рекомендуются некоторые схемы расчета состава сплава, как показано в таблице 2. Элементы в скобках составляли ≤0,01 вес.%, Что можно считать тривиальным.
Таблица 2 Рекомендуемые составы в соответствии с заданными свойствами UTS и EC на основе модели P2CТаблица 2 показывает, что при целевом увеличении UTS с 550 до 700 МПа рекомендуемые сплавы по модели P2C изменяются с серии Cu – Fe – P сплава (1 #) к сплаву серии Cu – Ni – Si (2–5 #).Для сплавов 2–5 # UTS сильно коррелирует с содержанием Ni; но нет очевидного изменения в других легирующих элементах, так как те <0,01 мас.% считаются примесными элементами. Мы обнаружили в литературе четыре экспериментальных сплава, которые очень близки к пяти разработанным сплавам в таблице 2. Их свойства и состав также перечислены в таблице 3. Лучшие значения UTS и EC для Cu – 2.35Fe – 0.10Cr – 0.03P Сплав 29 после гомогенизации, горячей прокатки, обработки раствором, холодной прокатки и старения составляют 495 МПа и 58.0% МАКО соответственно. Его состав очень похож на сплав 1 # в таблице 2. UTS очень близок (разница всего 5 МПа), а EC на 8,0% IACS выше, чем у сплава 1 #. Содержание Ni, Si и Mg во втором, 30 третьем 31 и четвертом 32 сплавах в таблице 3 аналогично содержанию 2 #, 3 # и 4 # в таблице 2. Измеренные ЕС второго сплава на 4,1% ниже IACS, чем целевое значение ЕС сплава 2 #, а измеренное значение UTS на 120 МПа выше, чем у сплава 2 # (21.8%). Экспериментальная ЭК третьего сплава на 6,0% (12,0%) по IACS ниже, чем у сплавов 3 # и 4 #, а экспериментальная UTS на 30 МПа выше, чем у сплава 3 # и на 20 МПа ниже, чем у сплава 3 #. 4 # сплав. Состав сплава Cu – 4.00Ni – 0.95Si – 0.02Cr – 0.02P 32 в таблице 3 аналогичен составу сплава 5 # в таблице 2, но их свойства существенно отличаются. Измеренное значение EC этого сплава на 15,0% ниже, чем у сплава 5 #, а измеренное значение UTS на 130 МПа выше, чем у сплава 5 # (18,6%).
Таблица 3 Сплавы с составами, аналогичными перечисленным в Таблице 2, и их свойства, описанные в литературеВидно, что две модели BP NN (модель C2P и модель P2C), обученные с помощью одного и того же набора данных, отличаются по эффективности обучения и прогнозированию способность. Модель C2P может получить удовлетворительные результаты прогнозирования производительности, а результаты расчета композиции, полученные с помощью модели P2C, менее надежны. В машинном обучении, когда количество обучающих выборок постоянно, предсказательная способность модели уменьшается с увеличением размерности предсказания (количества выходных переменных). 33 Мы предполагаем, что основная причина такой разницы заключается в разной топологии двух моделей. Модель C2P использует более десяти независимых переменных для подбора двух зависимых переменных, что является «подгонкой уменьшения размерности». Обучение модели легко сходится. Модель P2C соответствует более чем десяти зависимым переменным только с двумя независимыми переменными, что является «подгонкой увеличения размера». Сходимость модели плохая, и она склонна к переобучению. В большинстве случаев сплавы с высокими эксплуатационными характеристиками являются многокомпонентными.В результате использование модели P2C для обратного проектирования композиции легко сталкивается с упомянутым выше «приспособлением для увеличения размера», и, следовательно, трудно получить удовлетворительные композиции. Это могло быть важной причиной медленного прогресса в обратном проектировании составов сплавов. Чтобы решить эту проблему, необходимо разработать надежный подход к композиционному дизайну, чтобы быстро и точно спроектировать разумный сплав с заданными свойствами.
Построение и применение объектно-ориентированной MLDS
С этой целью мы предлагаем следующую систему проектирования с машинным обучением (MLDS), блок-схема которой показана на рис.3. Основная идея заключается в высокой надежности модели C2P и низкой надежности модели P2C, мы используем модель C2P для оценки схем расчета состава сплава, представленных моделью P2C, и установления критерия разумности расчета состава (т. Е. относительная ошибка между целевой производительностью и прогнозируемой производительностью). Путем повторения проб и ошибок системы MLDS можно автоматически и быстро проверить более разумную схему расчета состава сплава. Вот конкретные методы:
Рис.3Блок-схема системы проектирования машинного обучения (MLDS) для быстрого и точного композиционного дизайна
Во-первых, BP NN создает две модели C2P и P2C. Во-вторых, схема расчета начального состава сплава выполняется моделью P2C на основе заданных свойств. В-третьих, предыдущие схемы вводятся в модель C2P с более высокой надежностью для получения более точного прогноза. В-четвертых, прогнозируемые свойства сравниваются с целевыми и получается ошибка. По ошибке будет проводиться либо выбор схемы расчета композиции, либо переобучение модели.Если все ошибки между прогнозируемыми и целевыми значениями превышают заданный порог, модель P2C будет повторно обучаться до тех пор, пока не будет исключена разумная схема расчета состава сплава. MLDS разделена на три подсистемы, включая обучение модели, композиционный дизайн и прогноз свойств. Три подсистемы взаимосвязаны и образуют замкнутую систему.
Чтобы сравнить характеристики MLDS и только модели P2C по композиционному дизайну, мы установили целевые свойства UTS на 750 МПа и EC на 50.0% МАКО. В общей сложности 20 возможных сплавов были разработаны MLDS с порогом ошибки 10% и моделью P2C и перечислены в Таблице S2.
Концентрация основных компонентов (Ni, Si, Zn и Cr) в разработанных сплавах представлена на рис. 4. Колебания Ni, Si, Zn и Cr сплавов, разработанных по модели P2C, значительны, в то время как колебания сплавов, разработанных MLDS, небольшие и стабильные. Как показано в Таблице S2, средние концентрации Ni, Si, Zn и Cr в сплавах, разработанных MLDS, равны 2.29 мас.%, 0,51 мас.%, 0,25 мас.% И 0,06 мас.% Соответственно. В то время как сплавы, разработанные с помощью модели P2C, составляют 2,46 вес.%, 0,55 вес.%, 0,30 вес.% И 0,12 вес.% Соответственно, что на 0,17 вес.%, 0,04 вес.%, 0,05 вес.% И 0,06 вес.% Выше, по сравнению с со сплавами, разработанными МЛДС. Кроме того, с точки зрения стандартного отклонения, максимального и минимального значений прогнозов (т.е. диапазона колебаний композиции) MLDS работает намного лучше, чем модель P2C. Приведенные выше результаты показывают, что MLDS имеет более высокую эффективность и надежность при решении проблемы композиционного дизайна для заданных свойств.
Рис. 4Основные компоненты, a Ni, b Si, c Zn и d Cr, из сплавов, разработанных моделью P2C и MLDS с целевыми свойствами UTS 750 МПа и EC 50,0% IACS
Теперь мы устанавливаем целевое ЕС 50,0% IACS и UTS 600–950 МПа и проводим MLDS с порогом ошибки 10,0%. Схемы расчета состава сплава показаны в Таблице 4. 34,35,36 С увеличением целевого значения UTS концентрации элементов Ni, Si и Cr становятся больше.Фактически, элементы Ni, Si и Cr являются хорошо известными упрочняющими элементами для медных сплавов и могут образовывать упрочняющие фазы, такие как Ni 2 Si и Cr 3 Si, что приводит к значительному увеличению UTS. 2,26 Основная роль следовых элементов Zn и Sn заключается в увеличении энергии дефектов упаковки матрицы, что улучшает UTS, но отрицательно влияет на EC. 1 Основная роль микроэлементов Mg и P заключается в очистке расплава и повышении текучести расплава во время плавления и литья. 32,35 Также показано, что добавление P может способствовать выделению упрочняющих фаз и частиц Cr, 34,35 и, таким образом, может усиливать упрочняющий эффект Ni, Si и Cr. Это также улучшает ЕС сплава. Следовательно, в сплавах 5 # –8 # с увеличением концентрации упрочняющих элементов, таких как Ni, Si и Cr, P, также значительно увеличивается прочность. Таким образом, в таблице 4 Ni, Si и Cr являются основными упрочняющими элементами, в то время как Mg, P, Zn и Sn являются вспомогательными элементами, которые используются для баланса между механическими свойствами и электропроводностью сплава. .В таблице 4 элемент Fe не присутствует в расчетном составе сплава 1 # –8 #, поскольку упрочняющий эффект Fe не так заметен, как Ni, Si и Cr. Хотя добавление большего количества Fe может улучшить UTS медного сплава, чрезмерное добавление Fe будет иметь отрицательное влияние на ЕС сплава. Фактически, текущая серия сплавов Cu – Fe – P имеет низкий UTS и высокий EC.
Таблица 4 Результаты расчета и проверка состава высокопрочных и многоэлементных сложных медных сплавов с высокой электропроводностьюКак показано в Таблице 4, содержание Ni, Si и Cr как основных упрочняющих элементов, значительно увеличивается с увеличением целевого UTS в медных сплавах, разработанных по модели MLDS, а массовое отношение Ni / Si остается между 3.5 и 5.0. Эти результаты согласуются с основными принципами проектирования медных сплавов на основе металлургии. 37,38 С другой стороны, когда содержание упрочняющих элементов, таких как Ni, Si и Cr, значительно увеличивается, неизбежно ухудшается ЕС медных сплавов. Для решения этого противоречия схема, рекомендованная MLDS, заключается в увеличении концентрации элемента P, поскольку следовой элемент P может очищать матрицу и способствовать выделению второй фазы сплавов Cu – Ni – Si, что способствует улучшению ЭК сплавов Cu – Ni – Si. 39,40 Таким образом, результаты, полученные с использованием модели MLDS для расчета состава сплава, соответствуют основным принципам металлургии и дополнительно подтверждают возможность расчета состава сплава методом MLDS на основе машинного обучения.
Три сплава с аналогичными основными компонентами, собранные в литературе, перечислены в таблице 4. Условиями их обработки являются обработка раствора, деформация и старение. И они отсутствуют в нашей исходной базе данных, показанной на рис. 6. UTS и EC трех сплавов в таблице 4 очень близки к целевым свойствам, которые мы установили для MLDS, демонстрируя надежность MLDS и рациональность результатов проектирования.
В частности, концентрация Ni и Si в сплаве Cu – 1,37Ni – 0,28Si – 0,02Mg – 0,04Zn – 0,04Sn – 0,03P 34 аналогична концентрации Ni и Si в сплаве 1 #, разработанном MLDS. Его UTS очень близок к UTS сплава 1 #, а его EC на 3,0% IACS выше, чем у сплава 1 #. Основная причина заключается в том, что концентрация Zn в конструкционном сплаве высока и отсутствует элемент Р, который может улучшить ЕС сплава. Концентрация Ni и Si в Cu – 2.00Ni – 0.50Si – 0.30Cr 35 очень близка к концентрации Ni и Si в сплаве 3 #.Хотя сплав 3 # не содержит Cr, общее содержание элементов Mg, Zn, Sn и P составляет 0,31 мас.%, Что почти равно Cr (0,30 мас.%) Сплава в литературе. Хотя сплав 3 # не содержит значительного упрочняющего эффекта Cr и содержит больше Mg, Zn и Sn, которые также обладают эффектом упрочнения твердого раствора, UTS и EC сплава почти такие же, как целевые свойства сплава 3 #. в таблице 4. След P может способствовать выделению δ-Ni 2 Si, что способствует ЕС сплава 3 #.Для сплава Cu – 4.00Ni – 1.00Si – 0.10Mg 36 концентрация Ni немного ниже, чем у сплава 7 #, а концентрация Si и Mg немного выше, чем у сплава 7 #. В результате UTS выше, чем целевая UTS сплава 7 #, а ЕС ниже, чем целевая EC сплава 7 #. Кроме того, Cr и Si сплава 7 # могут образовывать упрочняющую фазу Cr 3 Si, что также благоприятно сказывается на ЭК. Микролегированный P также положительно влияет на EC, что приводит к большой разнице (относительная погрешность составляет 12.0%) между целевой ЭК и измеренной ЭК сплава 7 #.
Для дальнейшей проверки рациональности композиционного дизайна MLDS мы подготовили и измерили два из рекомендуемых сплавов в нашей собственной лаборатории, которые также перечислены в таблице 4. Cu – 2.20Ni – 0.42Si – 0.08Mg – 0.30Zn Сплав был использован для оценки модели C2P в таблице 1. Концентрация его основных компонентов близка к концентрации сплава 4 #. Ошибка между измеренными свойствами (UTS 720 МПа и ЕС 49,5% IACS) и целевыми свойствами сплава 4 # (UTS 750 МПа и ЕС 50.0% IACS) составляет <4,0%. Второй сплав Cu – 3.00Ni – 0.60Si – 0.16Zn – 0.15Cr – 0.03P был приготовлен в соответствии с составом сплава 5 # в таблице 4. Измеряли UTS и EC образцов, а результаты UTS как функцию времени старения показаны на рис. 5. Свойства оптимизируются после старения в течение 240 мин, что дает UTS 775 ± 10 МПа и ЕС 48,0 ± 0,5% IACS. Разница между измеренными и заданными свойствами составляет 25 МПа и 2,5% IACS (относительные погрешности 3,1% и 4,0%) соответственно.Измеренные свойства по разработанным составам хорошо согласуются с целевыми свойствами.
Рис. 5Предел прочности на разрыв как функция времени старения при 450 ° C сплава Cu – 3,00Ni – 0,60Si – 0,16Zn – 0,15Cr – 0,03 P
MLDS, показанный на рис. Настоящее исследование имеет высокую надежность при разработке состава сплава и может удовлетворить требования быстрого и точного расчета состава для заданных требований к свойствам (целевые свойства).
Таким образом, мы впервые предложили систему проектирования с машинным обучением, которая объединяет модель BP NN свойства-состава, которая разрабатывает составы, и модель BP NN состава-свойства для эффективного просмотра схем расчета состава сплава.Продемонстрирован быстрый и более точный расчет составов сплавов. MLDS предоставляет новый метод для ускорения открытия новых материалов.
Затем с помощью MLDS реализуется обратная конструкция восьми новых медных сплавов с UTS 600–950 МПа и ЕС 50,0% IACS, что может стать хорошими кандидатами для исследования и разработки медных сплавов с высокими эксплуатационными характеристиками для крупногабаритная выводная рамка интегральной схемы и высококачественные разъемы.
Наконец, рациональность композиционного дизайна MLDS подтверждается путем сбора трех сплавов из литературных источников и приготовления двух новых сплавов в нашей собственной лаборатории в соответствии с рекомендуемыми составами MLDS.Ошибки между измеренными свойствами и целевыми свойствами составляют <6%, что указывает на удовлетворительную композиционную конструктивную способность MLDS.
Эти списки в алфавитном порядке включают синонимы общепринятых названий минералов, произношение этого имени, происхождение имени и информация о местонахождении. Посетите наш расширена подборка картинок с минералами.
Другие алфавитные списки минеральных видов в Интернете
|
Сплав вольфрама и меди — Cu + W
Состоящий из порошка чистого вольфрама (W), взвешенного в матрице из меди и (Cu), эти сплавы легко обрабатываются и известны хорошей теплопроводностью и электропроводностью, низким тепловым расширением, немагнитностью, хорошими характеристиками в вакууме, и устойчивость к эрозии от дугового разряда . Таким образом, они часто используются в качестве электрических контактов, особенно в высоковольтных устройствах.Большинство размеров и форм могут быть поставлены в короткие сроки. Мы также можем изготовить детали из этих материалов по вашим требованиям.
Позвоните нам, чтобы задать вопрос или узнать цену , 1-630-325-1001, или заполните нашу контактную форму .
Преимущества:
- Высокое сопротивление дуги в сочетании с хорошей электропроводностью
- Высокая теплопроводность
- Низкое тепловое расширение
Заявки:
- Электроды для контактной сварки
- Дуговые контакты и вакуумные контакты в выключателях высокого / среднего напряжения или вакуумных выключателях
- Электроды для электроэрозионных станков
- Радиаторы и радиаторы для пассивного охлаждения электронных устройств
- Электронные упаковочные материалы
- Компоненты базовой радиостанции
- Модули IGBT для гибридно-электрических (HEV) и полностью электрических (EV) транспортных средств
Таблица химического состава и свойств медно-вольфрамового сплава
| Состав | Плотность г / куб.см | МАКО % | Твердость HB кгс / мм2 | Прочность на изгиб МПа |
| W55 Cu45 | 12.30 | 49 | 125 | – |
| W60 Cu40 | 12,75 | 47 | 140 | – |
| W65 Cu35 | 13,30 | 44 | 155 | – |
| W68 Cu32 | 13,93 | 43 | 165 | – |
| W70 Cu30 | 14,18 | 42 | 175 | 667 |
| W75 Cu25 | 14.50 | 38 | 195 | 706 |
| W80 Cu20 | 15,15 | 34 | 220 | 736 |
| W85 Cu15 | 15,90 | 30 | 240 | 765 |
| W90 Cu10 | 16,75 | 27 | 260 | 804 |